
1. Introduction
Cells in the human body are embedded in a three-dimensional (3D), confined microenvironment of tissue matrices and circulatory vessels.1 This extracellular matrix (ECM) transmits a complex mixture of chemical and physical cues to cells that are crucial for their physiology and function.2,3 Geometrical or mechanical constraints of the microenvironment are sensed by major mechanosensors: integrins in focal adhesions and/or cadherins in adherens junctions. Integrins serve as a link between the extracellular matrix and the cytoskeleton, facilitating the cell's ability to sense and respond to mechanical changes. When integrins are activated, they trigger a cascade of intracellular signaling, including the activation of focal adhesion kinase (FAK) and other proteins.4,5 This signaling propagates through various pathways, notably the RhoA/Rock/Myosin pathway, which is integral to cytoskeletal rearrangement and the regulation of cellular tension.6 Mechanical cues result in changes in the assembly of the cytoskeleton and actomyosin contractility, further transferring information about the mechanical properties of the ECM into the cell. In addition to these, the Piezo1 ion channel is another key player in mechanotransduction, activation of piezo1 leads to ion flux across the membrane, initiating downstream signaling events that influence various cellular functions,7 including the organization of cell architecture,8,9,10,11,12,13,14,15,16 movement,17,18,19,20,21,22,23,24,25,26,27,28 polarization,11,17,29,30,31,32,33,34 division,34,35,36,37,38,39 growth,40,41,42 and differentiation13,15,31,40,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60 at subcellular, cellular, and multicellular scale.
Since the extracellular environment is highly complex and heterogeneous, and the equally complex cellular response depends on a range of variables, it is highly desirable to develop in vitro techniques that isolate different aspects of mechanical cues and to examine how these mechanical cues regulate the phenotypic and functional cellular responses to mechanical stimuli. Much effort has been put into generating hydrogels that capture the essential characteristics of the natural ECM. However, culturing cells on top of 2D hydrogels or embedding cells in 3D hydrogels are often not conducive to establishing causal relationships between the cellular response and geometric and mechanical characteristics of the microenvironment. As a result, the effects of mechanical forces produced by the 3D confined microenvironment on different biological processes remain unclear. To address this problem, various groups have exploited lithographic tools to precisely control the sizes and shapes in different dimensions, known as micropatterning technology.15,19,61,62,63,64 This removes the heterogeneity inherent in both natural and synthetic hydrogel systems, and establishes a crucial opportunity for quantitative studies and fundamental research in cell biology (Fig. 1). Micropatterns refer to precise and controlled miniature patterns created on a substrate surface, often used to regulate the spatial distribution of cell-adhesive proteins. In the realms of biotechnology and cell biology, micropatterns serve as tools to guide the behavior and organization of cells, affecting their adhesion, morphology, proliferation, and differentiation by offering physical cues within their microenvironment. While often thought of in two dimensions, micropatterns can also include quasi-three-dimensional configurations like microwells, broadening their capacity to manipulate cellular microenvironments and interactions.

Fig. 1. The application of nano/micro patterning in fundamental cell biology. |
Here, we review key examples of micropatterning use in mechanosensation and transduction, and discuss recent discoveries in cell behavior and fate commitment in different dimensions. Various techniques, such as photolithography,65,66 soft lithography,67,68 microcontact printing,69,70 electron beam lithography,71,72 and dip-pen nanolithography,73,74 are employed to create micropatterns. However, the central aim of this review is not solely to summarize current micropatterning techniques, which has been extensively and excellently reviewed elsewhere.75,76 Instead, we approach this topic from a biologist's perspective, focusing specifically on recent progress that leverages micropatterns to regulate the cell and tissue functions in different dimensions. We believe this review will motivate researchers to explore new directions combining advanced methodologies with micropatterning, and expand our fundamental understanding in tissue self-organization in different scales and dimensions.
1.1. Dimensionality of micropatterns: from 2D to 3D
Although 2D micropatterns provide compelling insights into the interplay between physical constraints and cell function, emerging evidence indicates that cells cultured on 2D substrates significantly differ from 3D cultures in cell spreading, adhesion, division, and fate determination.8,15,26,46,64,77 In a 2D environment, cells adopt flattened morphologies and become polarized due to the asymmetric apical-basal distribution of adhesive proteins (Fig. 2a). It should be noted that polarization of migrating cells also occurs in a 3D microenvironment both in vivo and in vitro.78,79 However, it is difficult to isolate the effects of cell morphology (sizes and shapes) on cell behaviors from the chemical and physical properties of scaffolds in a 3D system. Nevertheless, moving from 2D to 3D micropatterning will uncover a deeper understanding of many fundamental questions in cell biology.
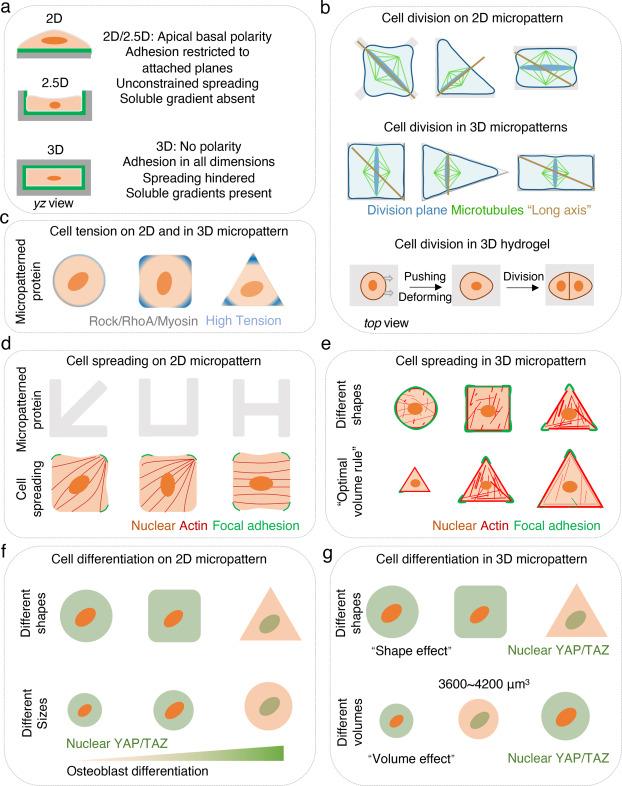
Fig. 2. Micropatterning provides a reproducible platform to investigate cell response to mechanical cues. |
One strategy to expand micropatterning dimensionality is to use pseudo-3D (or 2.5D) platforms, such as microwells39 or microgrooves,26,28 in which single or multiple cells are cultured in micron-sized cavities or gaps with defined geometry on a flat substrate. However, these pseudo-3D systems cannot fully eliminate biased cell polarization that is induced on a 2D flat substrate.
There are several ways to convert the pseudo-3D to entirely 3D microenvironments, with no biophysical and/or biochemical properties asymmetrically distributed across any sides of cells. For example, when seeding cells into the microwells, a liquid hydrogel produced from extracellular matrix proteins can be introduced to create a true 3D microenvironment.80 Alternatively, a hydrogel lid can be placed on top of the microwells to create a completely symmetrical microenvironment for the cells15,27,28,39 (Fig. 2a). Below, we compare different cellular responses between cells cultured on 2D and in 3D micropatterns.
1.2. Micropatterning-guided cell division from 2D to 3D
Cells cultured on 2D micropatterns become progressively flatter and divide aberrantly. When dividing in 3D, cells actively generate forces that push and deform the surrounding microenvironment, providing sufficient space for daughter cells37,38,80 (Fig. 2b). Interestingly, the cells’ physiological function can be reinstated when they are embedded in a 3D hydrogel. For example, the encapsulation of dedifferentiated chondrocytes restores their physiological phenotype, including cell shape and the expression of cartilaginous markers.81
The molecular mechanisms underlying cell division in 2D and 3D cultures exhibit significant differences due to the distinct interactions cells have with their environment. In 2D cultures, cells primarily interact with a flat surface, engaging with the substrate through receptors like integrins, which affects signaling pathways such as FAK and MAPK.82 In contrast, 3D cultures offer a more complex interaction with the surrounding matrix, allowing cells to establish multidirectional contacts that influence a variety of signaling pathways critical for cell division.83 This three-dimensional interaction promotes a more pronounced cellular polarity and an organization of the cytoskeleton in three dimensions, impacting the positioning of the centrosome and the orientation of the mitotic spindle during cell division.84
Micropatterning is a powerful method to study the orientation and position of the cell division plane, which is crucial for maintaining proper cell function. According to Hertwig's rule,85 cells always divide along the longest axis according to their shape.86 The orientation of cell spindles and DNA segregation are influenced by adhesive patterns on 2D micropatterns,11,34 thus determining the direction of mitosis.36 Notable exceptions to the “long-axis rule” emerge in 3D by placing a glass on top of the micropatterned microwells. Cells that are confined in these pseudo-3D microenvironments divide along the longest axis of symmetry rather than simply the longest axis39 (Fig. 2b). The orientation of cell division guides division symmetry, further determining cell fate allocations. Despite this growing understanding, we can address many outstanding questions using 3D micropatterns. For example, which signaling pathways are involved in determining the geometrical-determined division plane and mitotic spindle position? How does 3D cell geometry affect symmetrical or asymmetrical cell division? By understanding these questions, we can obtain new perspectives on mechanical regulation of cell division, in both normal and diseased tissue.
1.3. Micropatterning-guided cell tension and cell spreading from 2D to 3D
To observe the relationship between the geometrical signal and cell tension and the organization of cell structural elements, cells plated on micropatterned ECM islands are allowed to spread until they occupy the size and shape of the underlying adhesive area.41,42,63 Cells spread via pre-deposited adhesive proteins and are influenced by the size, shape, stiffness, and dimensionality of the substrate.
At the single-cell level and collectively, cell tension is consistently highest at the corners or edges of micropatterns, both in 2D and 3D10,18,23,87,88,89,90 (Fig. 2c). In 2D, the adhesive geometry of cells plays an important role in cell tension enrichment and cell spreading.13,43,44,45,91 Single cells plated on 2D micropatterns interact with the ECM through integrins and the formation of focal adhesions (FAs).12,13,14 This occurs predominantly at the cell periphery (circular)13 or cell corners (square) and is regulated by the micropattern shape9,10,12,14 (Fig. 2d). Single cells on round islands generate tensile forces along the circumference without a preferred direction, whereas cells on square islands tend to exhibit the highest tension at the corners.87,90 FAs are associated with the termini of contractile actin and myosin filaments, which generates tension in response to external and internal traction forces.10,12,14 Consequently, cells develop their stress fiber and tension distributions according to the geometry of the adhesive area.10,18,23,24,87,89,90 The most prominent contractile tension is generated along cell edges, particularly the concave edge of a non-adhesive region, where the stress fibers are anchored to ECM by FAs.9,10,12 Similarly, multiple cells on micropatterns also demonstrate the strongest stress fibers and contractility in the cells located at the edges.23,92 Through micro- and nano-patterning, researchers discovered that cell is affected by spatial constraints and adhesion. Although both approaches yield similar cell morphologies, they initiate distinct signaling pathways and influence cellular functions differently: nanopatterning via nano-grooves amplifies intracellular force and fosters osteogenic differentiation through the RhoA/ROCK pathway, while micropatterning through micro-grooves induces pseudopodia formation.93 Overall, cells sense the geometrical constraints produced by micropatterned ECM causing cytoskeletal filament reorganizations and actomyosin contractility transformation.
In contrast to cells cultured in 2D, cells that are encapsulated in a 3D hydrogel or settled in 3D microwells spread by pulling on the matrix and recruiting adhesive proteins to degrade their surrounding environment. MSCs demonstrate differential stiffness-related spreading tendencies based on their dimensionality. Specifically, in 2D substrates, MSCs expand with greater stiffness, however in 3D, MSCs expand with reduced stiffness. Cells in 3D may be unable to pull and deform the surrounding environment to build sufficient tension for spreading when stiffness is too high.8,16 Dimensionality also affects a cell's ability to sense spreading size and volume. In 2D micropatterns, increased spreading consistently occurs alongside higher cellular tension due to more pronounced FAs, more actin stress fibers, and increased YAP/TAZ activity.94 However, in 3D micropatterns, spreading may follow an “optimal volume rule”. When cells are confined in 3D microwells, smaller cells up to approximately ∼3600 μm3 have more F-actin polymerization, focal adhesion formation, and YAP/TAZ nuclear localization regardless of cell shape (Fig. 2e).8,15
1.4. Micropatterning-guided cell differentiation from 2D to 3D
Cell fate can be influenced by cell geometry at the single cell level. Changes to the cell spread range can induce shifts between apoptosis, growth, and differentiation.41,43,95 Single MSCs select different lineages while growing on varying 2D micropattern sizes and shapes.43,44 MSCs constrained on small ECM-coated islands spread poorly, with dis-organized actin cytoskeleton, and fewer focal adhesions (FAs), and the majority underwent adipogenic lineage.13,43 Similar MSC differentiation patterns occur when cells are constrained on low aspect ratio rectangles or convex pentagons.44 In contrast, MSCs culture on larger patterns, high aspect ratio rectangles or concave pentagons, exhibit strong stress fibers, present FAs along the periphery of the islands and nuclear YAP/TAZ localization, and show a preference for osteogenesis13,43,44 (Fig. 2f). While the complete understanding of the mechanisms behind Mesenchymal Stem Cell (MSC) lineage commitment remains elusive, there is established evidence indicating that cell geometry significantly influences this process. This influence is, in part, mediated through the Rho/ROCK signaling pathway, which plays a critical role in modulating cytoskeletal tension.43,44 A similar mechanism of cytoskeletal organization was identified in human keratinocytes, regulating stem cell differentiation under ECM-geometry confined conditions.45
In 2D micropatterns, cell spreading area, sizes and shapes dramatically influence hMSCs fate. In 3D, cell volume regulates hMSCs fate commitment.57,58,59 Rather than controlling cell morphology using micropatterns, cell shape and volume expansion in 3D may be achieved by modulating the properties of hydrogels. In contrast to hMSCs in 2D micropatterns, small volume, rounded cells embedded in hyaluronic acid-based hydrogels might differentiate efficiently into osteoblasts.57,59 However, MSCs encapsulated in viscoelastic hydrogels undergo osteogenic lineage that is associated with greater volume expansion.58,96 Further, the effects of cell shape on hMSC differentiation have been studied independent of the chemical and physical properties of hydrogels. As a result, round or elongated cells achieved similar differentiation potentials in this 3D context.59 Overall, these findings indicate that the mechanical properties of different hydrogels and cell morphologies may not directly dictate cell fate commitment, but rather impact how MSCs interact with ECM, further influencing mechanotransduction and stem cell fate allocations.16,97 3D micropatterning provides a unique opportunity to address how cell volume regulates differentiation. Single cell volume can be precisely controlled in hydrogel-made 3D microwells with a lid on top. There appears to be an optimal volume (∼3600 μm3) of hMSCs to differentiate into osteoblast, which is associated with well-established stress fibers and focal adhesions, YAP/TAZ nuclear localization, higher cell contractility, and nuclear accumulation of histone deacetylase 3 (HDAC3). Cell volumes that are too small (<3000 μm3) or too large (>6000 μm3) give rise to low tension and adipocyte differentiation15 (Fig. 2g).
1.5. Micropatterning-guided tissue patterning and self-organization
Micropatterning technology provides a platform to explore patterned differentiation of stem cells. Warmflash and colleagues first described the formation of spatially-organized germ layers along the radial axis of circular micropatterns following BMP4 treatment of human embryonic stem cells (hESCs).47 A series of follow-up studies examined how morphogen gradients were generated and their impact on patterned differentiation.48,49,50,51,52,53,54,55,56 Using high-throughput images and machine learning, Smith and colleagues predicted patterned formation of human-induced pluripotent stem-cells (hiPSCs) depending on cell density, cytoskeletal tension, and micropattern geometry.92 Furthermore, applying a microfluidic culture system allows spatiotemporally control over the morphogen sources (signaling center) and establishes dynamic concentration gradients, which artificially mimic the BMP4 source during the emergence of the primitive streak in vivo, opening up a new window to dissect geometry and biochemical gradients.98
Micropatterning was recently shown to model organogenesis, including neurogenesis and cardiogenesis. For example, by seeding hESC on micropatterns, neural organoids (termed neuruloids) were formed with correctly patterned lineages.99 These neuruloids may serve as powerful model systems to identify mechanisms underlying neurodegenerative diseases. Geometrical signals also affect human pluripotent stem cell differentiation after neural induction using a similar micropattern approach This study also determined that geometrical confinement generated by micropatterns induces patterned differentiation of neuroepithelial and neural plate border cells, recapitulating early neuroectoderm differentiation during in vivo100
hiPSCs seeded on micropatterns can be also used for mimicking cardiogenesis. Ma and colleagues found that hiPSCs in the center of micropatterned differentiated into cardiomyocytes after cardiac induction, whereas cells at the periphery differentiated into stromal cells, as indicated by smooth muscle marker staining (Fig. 3a).101
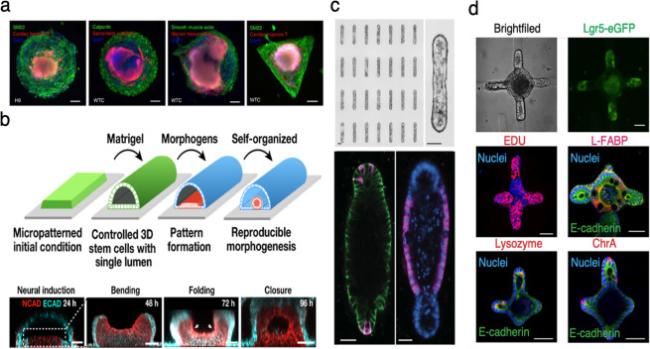
Fig. 3. Micropatterning-guided tissue patterning and self-organization |
More recently, micropatterning became a powerful technology to model tubulogenesis and neural tube morphogenesis in vitro with high throughput and reproducibility.104,105 By culturing cells in either 2D or 3D micropatterns and using extracellular matrix protein (for example Matrigel), the tissue is folded to form a tubular-like morphology. Impressively, this technology has recently been used to generate human neural tubes.102 A robust transition from the constrained 2D pattern into a 3D tubular morphogen with a single lumen may be observed followed by TGF-β and BMP4 treatment (Fig. 3b), which offers a new direction in the organoid field for developing a geometry-controlled 3D model system.102 Furthermore, Lutolf's group utilized bioengineered microwells to precisely control the initial geometry of intestinal organoids, which then formed deterministic patterns (Fig. 3c and d).103
1.6. Outlook
Micropatterned biology has become an essential sub-discipline helping to elucidate key phenomena in cell biology. The development of micropatterned systems reveals important insights into the impact of geometrical and physical cues from the microenvironment in 2D and 3D on cellular function, from intracellular organization to morphogenesis, and from single cells to multiple cells (Fig. 4). The ability of cells to convert mechanical forces through micropatterning into biochemical signals is important throughout life, guiding cell fate commitments during development or regeneration, and responding to injury through guided cell migration. Micropatterning technology also contributes to our understanding of the geometric role of ECM during numerous biological processes and it demonstrates its potential to integrate with other advanced approaches.102
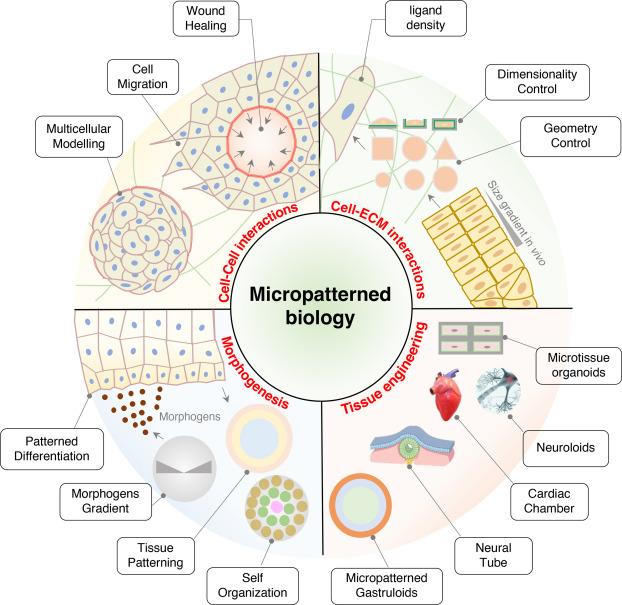
Fig. 4. The application of micropatterned biology. |
Micropatterning holds promise as a valuable tool in drug screening, given its profound influence on cell morphology. By dictating cell shape through strategic placement of micropatterned proteins, researchers can monitor the effects of pharmaceutical compounds on cells. Adverse drug reactions would likely disrupt the cells' predetermined morphological configurations, a change that could be utilized as a robust readout. This morphological alteration offers a direct and measurable marker of a drug's effect on cellular architecture and functionality. Employing micropatterning in this context could significantly improve the accuracy and speed with which compounds that negatively affect cell health and structure are identified.
Moreover, micropatterning offers a sophisticated approach to engineering organoids and tissue-like systems with diverse and precise shapes, enhancing the fidelity of in vitro models to replicate the complex structures of various tissues and organs.106,107 Consequently, researchers can examine cellular responses in conditions that closely mirror their natural environments. This enhanced simulation is invaluable for exploring cellular interactions within their microenvironment, their differentiation pathways, and their contributions to the tissue's or organ's overall functionality.23,108,109 Additionally, this approach enables scientists to analyze how the geometry of tissues and organs influences cell behaviour. For example, the spatial organization of cells can affect their proliferation, differentiation, and intercommunication—key elements in the development and functionality of tissues.103
Given the complexity of the native ECM, the key challenge of advanced biomaterial design will be to discover new materials that allow comprehensive integration of all required chemical and mechanical cues, while remaining amenable to microscale patterning and cell culturing.110 The integration of microfluidic devices and micropatterning technique would allow the creation of homeostatic environments or mimic blood vessels, allowing for even more complex tissues architectures.111,112 Micropatterned systems often provide a simplified representation, lacking the intricate interactions among various cell types and ECM components that characterize natural tissues. This simplification may restrict the relevance of insights gained from micropatterned systems to actual biological scenarios, particularly in the context of chronic diseases and extended tissue development. To mitigate this limitation, integrating more sophisticated and dynamic elements into micropatterned systems could be a viable approach. For instance, combining microfluidics with micropatterning could enable the dynamic control of chemical gradients and mechanical forces, more accurately simulating in vivo conditions.113,114 Additionally, employing hydrogels with adjustable mechanical properties or incorporating degradable materials that evolve could offer another avenue to replicate the dynamic environment,115,116,117 adding a more authentic temporal aspect to studies of tissue development and disease progression.
Investigating cell behavior in extended cultures through micropatterned systems offers critical insights into phenomena such as the progression of chronic diseases and the nuances of prolonged tissue development. Nonetheless, traditional 2D micropatterning faces challenges for sustained cell culture endeavors. As cells proliferate within micropatterned confines, their secretion of the ECM can gradually obscure the micropatterns, compromising the intended spatial limitations. To ensure persistent cell confinement over lengthy durations, employing microwells with defined geometries emerges as a viable strategy. For example, we have previously utilized this strategy to conduct in-depth studies on gap closure during wound healing and migration from microtissues in extended culture periods.23,108 Such microwells can establish a robust physical barrier, fostering continuous spatial restriction and more accurately simulating the cellular microenvironment observed in vivo.
Given the rapid development of high-throughput and high-resolution biological assays, human ESC gastruloid and mini-gut tubes generated from micropatterns have been analyzed by single-cell RNA sequencing (scRNA-seq) to explore cell heterogeneity and identify rare cell types.112,118 Future investigations may leverage micropatterning combined with spatial-omics,119,120 synthetic biology, optogenetics, quantitative live cell imaging, and genetic modification to delve deeper into self-organization principles, morphogenetic processes, molecular mechanisms of organ regeneration, and patterning plasticity, potentially adopting an evo-devo perspective. Emerging automated and standardized imaging platforms that integrate micropatterning with artificial intelligence could facilitate the acquisition of extensive multi-parametric datasets, enabling high-throughput studies on the impacts of various biophysical and biochemical signals on cell and tissue functions. Additionally, developing micropatterning platforms with stimuli-responsive materials could create dynamic environments that more closely mimic organogenesis and tissue development processes. Such advancements, validated across diverse in vitro and in vivo models, promise to enhance the translation of micropatterned biology to clinical settings, offering deeper insights into disease development and cell-cell interactions during development.
Ethical approval
This study does not contain any studies with human or animal subjects performed by any of the authors.
CRediT authorship contribution statement
Xinyu Hu: Writing - review & editing. Min Bao: Conceptualization, Funding acquisition, Supervision, Writing - original draft, Writing - review & editing.
Declaration of competing interest
The authors declare the following financial interests/personal relationships which may be considered as potential competing interests: Min bao reports financial support was provided by Oujiang Laboratory. Min bao reports a relationship with Oujiang Laboratory that includes: employment. If there are other authors, they declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgements
M.B.'s work in the lab is supported by start-up funding (grant OJQD2022015) from Oujiang Laboratory and the National Natural Science Foundation of China (grant 82301873).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}