
1. Introduction
Dendritic cells (DCs), as coined by Steinmann and Cohn, are a heterogenous group of cells classed as antigen presenting cells (APCs) as they possess a specialized capacity for engulfing and processing foreign antigens that are then presented to lymphocytes to initiate secondary immune responses.1,2 They are morphologically different from macrophages, immune cells that also have antigen presenting capacity, due to their distinct dendrites.3,4 DC's importance stems from the well acknowledged fact that they are two orders of magnitude more potent than all other APCs in engulfing and processing antigens for initiation of the adaptive immune response, as well as the induction of immunological tolerance, through communication with lymphocytes.5,6 During the adaptive immune response, DCs play the role of initiating antigen specific activation of lymphocytes whereas in immunological tolerance, they assist in guiding the immune system to avoid generation of autoimmune responses to self antigen.7,8
Within peripheral tissues in steady state, DCs exist as immature (iDCs) or semi-mature, having a higher capacity for antigen uptake and low expression of costimulatory molecules such as CD40, CD80, and CD86.9,10,11 Using specialized pattern recognition receptors such as toll-like receptors (TLR), nucleotide oligomerization domain-like receptors (NLRs), retinoic acid-inducible gene-I (RIG-I)-like receptors (RLRs), C-type lectin receptors (CLRs), and mannose-like receptors, iDCs scavenge, identify and phagocytose invading pathogens.4,12,13,14 Upon phagocytosis, iDCs process antigens and develop to form mature DCs (mDCs) which have an increased expression of major histocompatibility complexes (MHC) and costimulatory molecules.15 mDCs release large amounts of pro-inflammatory cytokines, signaling other innate immune cells in order to limit the spread of infectious agents.4 They then migrate from peripheral tissues into lymphoid organs where they present antigens on MHC proteins to activate antigen-specific T cells, thereby initiating the adaptive immune response.11,16 mDCs exhibit an increased expression of chemokine receptors, such as CCR7, which are needed to guide their migration to the nearest lymph node.17,18 It is important to note that DCs may prime qualitatively different kinds of effector T cell responses depending on the type of immunogenic stimulus they receive.19
1.1. DC ontogeny
Like all other leukocytes, DCs originate from hematopoietic stem cells within bone marrow.3 Increasing evidence of the heterogeneity of DC functions points to the fact that DCs exist as many distinct subtypes.20 Based on the developmental pathways from which they stem, DCs may be classified through their lineage as lymphoid- and myeloid-derived DCs.16,20,21 Lymphoid-derived DCs originate from the common lymphoid progenitor and are found residing within T cell-rich areas of lymphoid tissues where they regulate immune tolerance.16,22,23 Myeloid-derived DCs, on the other hand, originate from the common myeloid progenitor and are mainly found in non-lymphoid tissues.24 These two DC subsets are distinguishable by their expression of CD8α, with lymphoid DCs having an increased expression of this marker.25,26 Another classification of DCs divides them into conventional (or classical) DCs and non-conventional DCs.21,27 Conventional DCs are cells that have already developed a certain form and function and are found in steady state within tissues.21,28 They are further divided into conventional DC1, a highly specialized subset for presenting exogenous antigens on major histocompatibility complex class I (MHC-I) to activate CD8 T cells, and conventional DC2, which are highly specialized in MHC class II-mediated antigen presentation for activation of CD4 T cells.29,30,31 Non-conventional DCs, on the other hand, refer to those that develop from DC precursors (pre-DCs) and are formed in response to inflammatory stimuli.21,28 Non-conventional DCs are further subdivided into plasmacytoid DCs which are mainly responsible for the production of type I interferons in response to viral infections6,32 and monocyte-derived DCs that develop from monocytes recruited from circulating blood during inflammation.33 Other specialized DC subsets exist in specific tissue including the skin (Langerhans cells and dermal DCs), lungs, kidney, liver, and intestinal tract.21
1.2. DC immunomechanobiology and the tumor microenvironment
Given their potent capacity to present antigens, thereby initiating the T cell response, DCs play a crucial role in instigating and sustaining anti-tumor immunity. Unfortunately in the case of cancer, DCs undergo a state of dysfunction, and there is increasing evidence indicating that it is primarily attributed to the immunosuppressive characteristics of the tumor microenvironment (TME).34 The immunosuppressive nature of the TME is influenced by numerous factors and characteristics, with a notable contribution from its distinctive mechanical features.
In the TME, there is a shift in balance of fluid between blood, interstitium and lymph that gives rise to interstitial fluid pressure and this is due to a combination of hypoxia-driven angiogenesis of leaky vessels and remodeling of the ECM at tumor boundaries.35,36,37 As a result, cells within the TME are exposed to increased fluid shear forces. As proven by several in vitro studies, fluid shear forces activate fibroblasts into myofibroblasts within the TME and the contractile phenotype aligns collagen fibers within matrices in a long range manner.38 Aligned matrices also mechanically maintain myofibroblast phenotypes, and hence a feed forward signal is present.39 The level of myofibroblastic-like activity and number of myofibroblasts is dependent on the intensity of stimuli, amplitude of fluid shear forces or extent of alignment. Similarly cancer cells also align matrices through contractile activities, although it has not been shown if this is enhanced due to an exposure to increased shear forces.35 Notably, increased matrix stiffness contributes to tumor progression.40,41 There are additionally cancer associated fibroblasts, adipocytes, cancer associated adipocytes, cancer associated macrophages, stem cells, and a plethora of immune cells that have not been systematically studied. Regardless, within the TME, both ECM composition42 and stiffness43,44 changes due to concerted contribution of several of these cell types participating in matrix remodeling activities. Interstitial flow in TME is heterogeneous36 and difficult to measure, and by virtue of their interrelationship there exists mechanical heterogeneity in the TME.
The unique mechanical signatures within the TME imparts diverse forms of biomechanical forces that may collectively contribute to the altered function of immune cells, including DCs. Such biomechanical forces can influence the behavior, and immunological functions of DCs.45,46 DCs capability of sensing and responding to mechanical forces during their interactions with their microenvironment and with other cells is manifested through the related works compiled in this review. Correspondingly, DCs also exert mechanical forces which can influence ECM organization47 as well as behavior of other cells. Our understanding of DC mechanobiology is essential as it provides insights into mechanisms by which DC functions may or may not be altered by biomechanical cues within the TME, thereby fueling the development of de novo therapeutic approaches. In this review, we discuss the current knowledge and understanding of DC mechanobiology through two main lenses: DCs as sensors of mechanical forces and DCs as providers of mechanical forces. An overview of these stimuli and their effects is illustrated in Fig. 1.
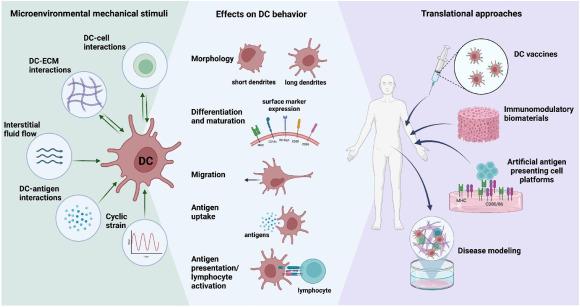
Fig. 1. Schematic summary of DC mechanobiology and its clinical applications. |
2. Dendritic cells as sensors of mechanical forces
DCs are exposed to various mechanical forces throughout their lifespan. These mechanical forces are present from the start of their journey as immature DCs performing surveillance in the TME, through their maturation and trafficking across tissues en route to lymphoid organs, and ultimately as matured DCs homing to lymph nodes. The mechanical forces may come from their interactions with other cells, from changes in biomechanical properties and topographical properties of the tumor ECM, and from elevated shear stresses as a result of the increased fluid pressure in the ECM and vascular network of tumors. Such mechanical stimuli may affect DC differentiation, maturation, morphology, migration, phagocytic function, as well their antigen presentation and activation of T and B cells. Here, we discuss the collective research efforts unraveling how these mechanical cues influence DCs and how these findings can be used to fuel immunomodulatory therapeutic strategies.
2.1. Substrate dimensionality affects DC morphology
DCs possess a unique morphology with long, branched projections called dendrites that extend from their cell body. Within the dendritic extensions of DCs are mechanosensitive adhesion structures called podosomes consisting of an actin rich core surrounded by integrins, adaptor proteins, and an actin network filament.48,49 Podosomes are involved in numerous functions including cell matrix adhesion, mechanosensing, and cell migration.49 Individual podosomes probe their microenvironment by undergoing cycles of repetitive protrusion and retraction.50 In podosome clusters, these oscillations are synchronized in a wave-like manner.51,52 Gong et al. have shown that DC podosomes show a growth in these oscillations when polymerization-driven protrusion and signaling-associated myosin contraction occur at similar rates.53 Additionally, the wave-like coordination of the oscillations are driven by diffusion of G-actin monomers.53 The actin polymerization in the podosome core which results in protrusive forces that cause DC dendrite formation49 has been shown to be controlled by the Rho family of GTPases, including RhoA, Rac1 and Cdc42.54 Collectively, these contribute to DC mechanosensation of diverse biomechanical cues within their microenvironment, influencing their morphological characteristics and immune functionalities.
Due to its constant remodeling and reorganization which causes its varying degrees of localized stiffness and topography, the ECM is a source of biomechanical cues which can influence cellular behavior.55 The ECM of cancer tissue, for one, exhibits several distinctive features and signatures compared to ECM of normal tissue, enabling it to deliver distinct mechanical cues to cells. More specifically, tumor ECM is characterized by increased collagen deposition and cross-linking mediated by lysyl oxidase enzyme.56,57 Increased ECM deposition, as well as crosslinking, elevates the stiffness of tumor ECM, providing mechanical cues that affect DC morphology. In vivo, biomechanical modulation of DC morphology by altered ECM properties has been demonstrated in the medical condition, acromegaly. In acromegaly, patients overproduce growth hormone resulting in somatic disfigurement,58 including enlargement of the skin, thereby altering the mechanical properties of the dermis ECM.59 Quatresooz et al. showed that the numbers of Factor XIIIa-positive dermal dendrocytes, a specific population of dermal DCs, is significantly reduced in the superficial dermis of the acromegaly ECM causing an alteration in DC morphology towards a more plump and globular shape with only rare and small dendrites.59
Although not many studies exist in regards to the study of DC morphology within different mechanical microenvironments in vivo, there are reported uses of different in vitro systems to study how DC morphology is affected by mechanical stimuli of their microenvironments, in this instance, dimensionality. When cultured in 3D collagen type I scaffolds, murine bone marrow derived DCs (BMDC) exhibited an irregular shape with short and thick dendrites (Fig. 2A(i)) compared to those cultured in 2D, which presented a corona-like-radiating appearance with long and slim dendrites (Fig. 2A(ii)), indicating that the 3D geometry of the collagen scaffolds induced a change in DC morphology.60 The collagen scaffold used by Fang et al. refers to scaffold generated by dialyzing collagen solution, lyophilizing, and crosslinking to form porous collagen pellets (Fig. 2B(i)).61 This is different from fibrillar 3D collagen matrices, also discussed in this review, which are hydrogels used to mimic the structure of in vivo ECM (Fig. 2B(ii)). Despite the differences in the nature of the microarchitecture of collagen scaffolds vs matrices, we have previously made similar observations of morphological differences of DCs cultured within 3D collagen type I matrices of different densities in comparison to being cultured on 2D substrates (Fig. 2C). In our study, we examined the morphology of human THP-1 derived DCs in 2D and 3D culture systems of different matrix density from the tissue-centric perspective and the cell-centric perspective (Fig. 2D).62 From the tissue-centric perspective, where DCs are differentiated οn the scaffolds itself, we observed that DCs showed more pronounced dendritic protrusions on 2D compared to 3D, where shorter dendrites were observed62 (Fig. 2C). This tissue-centric perspective highlights how the specific microenvironments affect DC morphology. In the cell-centric perspective, where DCs are pre-differentiated from monocytes on 2D tissue culture polystyrene and transferred to a new microenvironment, DCs exhibited an increased cell area and elongated morphology in 3D matrix but showed no change in cell area and shape for 2D (Fig. 2C).62 Incidentally, the cell-centric perspective reflects the clinical application of matured and activated DCs for adoptive cell transfer immunotherapies.63,64
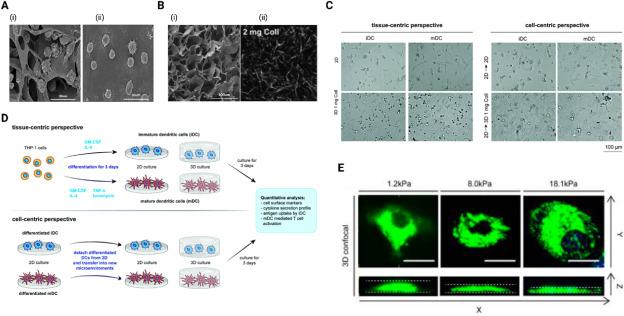
Fig. 2. Morphological differences of DCs in different in vitro cell culture systems: (A) Scanning electron microscope (SEM) images of DCs differentiated on (i) 3D collagen scaffold and (ii) 2D culture.60 (B) Representative image of (i) 3D collagen scaffold visualized using scanning electron microscopy (SEM)60 and (ii) 2 mg/ml 3D collagen matrices visualized using confocal laser scanning microscopy.62 (C) Representative images of DC morphology in 2D and 3D microenvironments from the tissue-centric and cell-centric perspectives.62 (D) Schematic summary of experimental setup from tissue-centric and cell-centric perspectives.62(E) Representative confocal microscope images of DCs showing distinctively different morphology under different levels of mechanical confinement in PDMS gel confiners.65 |
Morphological changes in DCs can also be induced through interaction with hyaluronic acid (HA) with our 3D collagen matrices. This holds significance due to the degradation of HA into lower molecular weight forms within the tumor microenvironment (TME).66 We showed that low molecular weight (LMW) HA elongated DC morphology with heightened dendritic protrusions, while high molecular weight (HMW) HA induced a rounded morphology and increased in clustering of DCs.67 While various other biochemical signals from the ECM can alter morphology, we specifically highlight HA, not just for its relevance in the TME, but also because its signaling has been reported to be associated with mechanical responsiveness.68 Furthermore, since cell morphology is governed by the cytoskeletal arrangement,69 and it is acknowledged the microgravity exposure alters the cytoskeleton, we studied DCs cultured within 3D collagen matrices in simulated microgravity (using Random Positioning Machine (RPM)). Interestingly, the DCs exhibited no morphological differences compared to those cultured on ground (not exposed to microgravity,70), despite a body of evidence indicating that microgravity impairs immune functions.71
In the 3D ECM, cells also come under geometric confinement within pores created by the interlocking arrangement of collagen fibers and other proteins forming the ECM. In the ECM of TME where there is increased matrix deposition and crosslinking, there is generally a decrease in size of these pores, enabling the ECM to exert various compressive mechanical forces on cells as they move through the pores. Additionally, the uncontrolled growth of tumor cells in a small space, or the change in HA molecular weight, creates confinement, generating other compressive mechanical cues within the tumor.72 Murine BMDC morphology has been found to be modulated by external mechanical stiffness of confined space, with a distinctly increased cell height observed at low mechanical load (1.2 kPa) and more flattened morphology/increased cell surface area at higher mechanical loads of 8.0 and 18.1 kPa65(Fig. 2E).
Overall, these studies show that biomechanical stimuli, in terms of dimensionality and topology, can affect the morphology of DCs, in particular the shape and length of dendritic protrusions. This may, in turn, affect downstream DC functions as per the theory of form vs function observed in many cell types.73,74,75 As a cautionary however, from our microgravity work on immune cells, morphology does not completely reveal DC immune capabilities.
2.2. Mechanical force and substrate dimensionality influence on DC differentiation and maturation
One of the key downstream functions which holds utmost importance is the differentiation and maturation of DCs. DC differentiation is a complex process involving several stages that are modulated by various signals from the microenvironment, including mechanical stimuli. Differentiated DCs exist within tissues as iDCs, possessing specialized pathogen-associated molecular patterns or danger-associated molecular patterns that enables them to scavenge and recognize foreign antigens. Following recognition and antigen uptake, iDCs undergo maturation to form mDCs which possess upregulated expression of MHC proteins and costimulatory molecules. The potency of maturation, as measured by known DC maturation characteristics, has also been shown to be influenced by different microenvironment mechanical stimuli. Here, we discuss studies showing how various mechanical stimuli from the tissue microenvironment, including exogenously applied cyclic strains, shear stresses, and cell-to-cell as well as cell-to-ECM interactions (substrate dimensionality, substrate stiffness, and tissue stiffness) affect DC differentiation and maturation.
Cyclic strain is a ubiquitous mechanical force that affects cells, including DCs, and may differ in magnitude and frequency depending on the type of tissue. Cyclic strain may arise from the pulsatile nature of blood flow within blood vessels,76 peristaltic and segmental contractions within the intestinal system,77 repeated breathing action of the lungs,78 contraction and relaxation of muscles forming part of the musculoskeletal system, dilation of bladder wall during urination, and repeated stretching of the skin.79 Within the TME specifically, repeated stretching of ECM fibers by stromal and cancer cells perhaps during the constant remodeling of the TME may also generate cyclin strain which could modulate DCs. Lewis et al. have demonstrated that application of 1 Hz cyclic strain of magnitude 10% for 24 h adversely affected murine BMDC viability compared to no-strain controls, and dependent on various adhesive substrates used for culture (e.g., laminin, collagen, fibrinogen).80 Application of the 1 Hz cyclic strain of magnitude 3% for 1 h, on the other hand, did not affect BMDC viability and even led to an increased expression of major histocompatibility complex class II (MHC-II) and costimulatory molecules CD86 and CD40 with this observed effect being more pronounced in BMDC cultured on pre-coated adhesive substrates.80 In line with the increased expression of costimulatory molecules and MHC-II, 3% cyclic stain for 1 h also led to increased stimulation of allogenic T cell proliferation.80 Cytokine secretion was however not significantly affected by cyclic strain.80 In the study, while no rationale was provided for the magnitude or duration of applied cyclic strain, it is palpable that both these parameters have an effect on BMDCs and warrants further investigation, perhaps with magnitudes and durations that are physiologically relevant.
Shear stress is also another source of mechanical stimuli that affects DCs. Shear stress results from the flow of interstitial fluid through tissues, as well as blood or lymph flow through the vascular network. The TME is well-characterized by increased angiogenesis,81 and the newly formed blood vessels tend to be disorganized and leaky.82 As a result, abnormal amounts of fluid tend to leak out into the surrounding tissue. In normal tissues, leakage of fluid from blood vessels is an essential physiological process for delivering oxygen and essential nutrients and for maintaining tissue homeostasis83 and the lymphatic system is responsible for draining interstitial fluid.84 However, due to impaired lymphatic drainage in the TME, there may be flawed clearance of fluid resulting in fluid build up and elevated fluid pressure in the TME.37 In addition, the dense and fibrotic tumor ECM creates physical barriers to fluid movement within the TME. All these in addition to rapid growth of tumor cells within the confined space of the TME contribute to elevating the pressure of interstitial fluid within the TME. Heightened shear forces generated as a result of interstitial fluid pressure provide mechanical cues that also influence DC activation and maturation. Murine BMDCs, upon exposure to shear stress in a microfluidics model system (Fig. 5B), exhibited increased expression of MHC-I and CD86 compared to those under static conditions.46,85 Increased extracellular pressure, applied through placing the cell culture system in an airtight pressure-controlled container, has also been shown to lead to increased secretion of IL-12, IL-6, TNF-α, and IFN-γ and expression of CD80, CD86, CD83, and MHC-II, indicating that extracellular pressure may act as a stimulus for DC maturation.86,87 Cytokine-matured DCs were however found to be less responsive to extracellular pressure compared to LPS-matured DCs.87
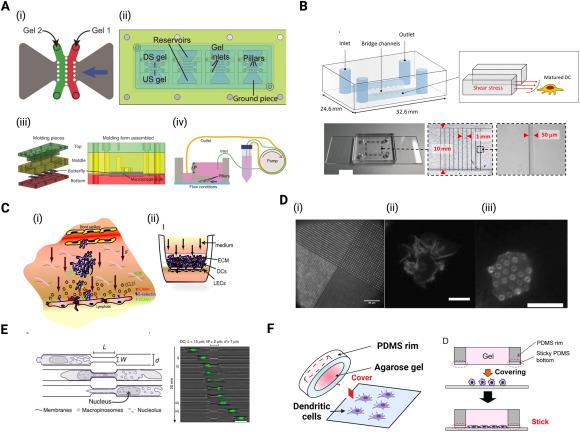
Fig. 5. Engineered in vitro systems to study effect of mechanical microenvironmental stimuli on DC migration dynamics (A) Microfluidic device used for 3D cell culture to study migratory behavior of MDA-MB-231 breast cancer cells in response to interstitial flow. The device contains (ii) 4 units of (i) two gels compartments upstream and downstream. (iii) The flow chamber is made up of a top, middle and bottom part defining the reservoirs and the ground piece creating gel channels. (iv) Schematic of the entire experimental setup for flow conditions93 (B) Schematic and actual photograph of microfluidic system used to expose DCs to shear stress.85 (C) (i) Schematic representation of transmural flow and DC migration from the ECM across lymphatic endothelium and (ii) in vitro model of the DC-lymphatic microenvironment showing lymphatic endothelial cells seeded on the bottom of a porous culture insert and ECM containing DCs through which medium flows at 0.1 or 1 μm/s, from the basal to the apical side.94 (D) (i) Micropatterned stamps used to produce microarrays by transfer of 488-conjugated fibronectin to PDMS coated coverslips for studying the effects of ligand geometry on DC migration. (ii) Representative images of DCs on (ii) PDMS-coated coverslips vs on (iii) microarrays.95 (E) Schematic representation of experimental setup used to study DC migration through narrow contractions and representative DC images stained with Hoechst (green).96 (F) Custom gel conifer designed to mimic in vivo geometric confinements for study of the effect of mechanical stiffness of narrow contractions on DC migration.65 |
Varying the substrate stiffness and also other physical surface properties presents different mechanical stimuli that differentially modulate DC differentiation and function. In a study in which human DCs were cultured on clinical grade titanium with different surface roughnesses and chemistries (Fig. 3A), it was observed that both smooth and rough surfaces promoted DC maturation compared to iDCs cultured on relatively smoother tissue culture dishes.88, 97, 98 However, surface roughness coupled with modification using carbonate and hydrocarbonate groups promoted the iDC phenotype.88 In addition, substrate stiffness has been shown to regulate the expression of β2 integrin but does not affect β1 integrin expression, with β2 integrin being 1.5 fold lower on human primary monocyte-derived dendritic cells derived iDCs conditioned on 12 kPa compared to iDCs conditioned on 2 or 50 kPa polyacrylamide (PAA) substrate.99 β2 integrins play a crucial role in formation of podosomes (Fig. 3B)89 and the loss of the β2 integrin cytoskeletal linkage has been shown to result in a transcriptome level reprogramming of DCs towards their mature, migratory phenotype.100 This suggests that substrate stiffness affects podosome formation and the adhesive behavior of iDCs.101 iDCs cultured on 2 kPa substrate also showed an increase in cell-cell adhesion and had an increased formation of clumps compared to 12 and 50 kPa substrates.99 Van den Dries et al. further reported that culturing DCs on flat (2D) and micropatterned (2.5D) substrates also dictates spatial organization and density of podosomes (Fig. 3C) and plays an important role in controlling DC activation and maturation.90 While on 2D surfaces iDCs cultured with Prostaglandin E2 (PGE2) causes rapid dissolution of podosomes, 2D surfaces with features (2.5D) inhibited PGE2-mediated RhoA activation resulting in impaired podosome dissolution.90 Podosomes are more prevalent in iDCs and their formation is lost upon maturation.91 PGE2, which is known to play a key role in DC maturation,102,103 has been shown to be responsible for mediating this loss of podosomes (Fig. 3D) during migration via myosin-II-mediated contractility downstream of RhoA and Rho-kinase activation.91 Following their maturation, mDCs cultured on 2 and 50 kPa PAA substrate upregulated the expression of CD83 but substrate stiffness showed no differences in CD86 and MHC-II expression.99 Chakraborty et al. also observed an elevated glycolytic rate and lactate production, increased expression of pro-inflammatory markers CD86, MHC-II and CD80 and upregulation of pro-inflammatory cytokines IL-1α, IL-1β, IL-6, IL-12, monocyte chemoattractant protein-1 (MCP-1), macrophage-inflammatory protein-2 (MIP-2), and TNF-α on BMDC conditioned on stiff (50 kPa) PDMS substrate.104 This sensing of mechanical stiffness by DC was found to be regulated by the Hippo-signaling pathway, YAP/TAZ as well as Ca2+ related ion channels including Piezo1.104
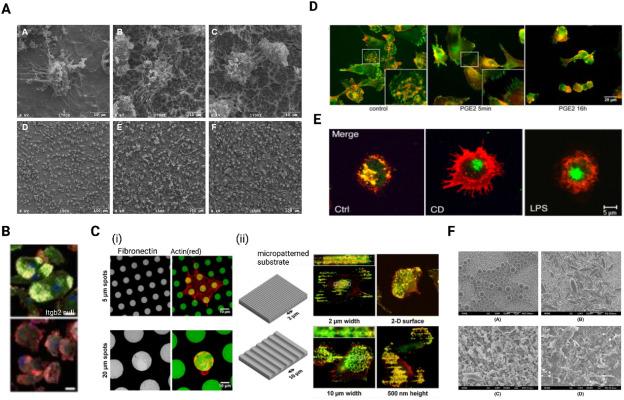
Fig. 3. (A) SEM images DCs on smooth (A, D) and rough titanium surfaces (B,C,E,F).88 (B) Wild type (WT) and β2-integrin (itgb2)-null DCs cultured on glass coverslips, fixed and stained for β2 integrin (green; FITC), F-actin (red; Alexa-Fluor-555) and vinculin (grey; Alexa-Fluor-633) showed loss of β2 integrin led to deficiency of podosome formation.89 (C) (i) DCs seeded on fibronectin/rIgG1-FITC (green) printed hydrogel and fixed and stained with phalloidin-Texas Red to visualize actin (red). Representative images of DC seeded on 5- and 20-μm dots shows adhesive fibronectin spots modulate spatial rearrangement of DC podosomes. (ii) Schematic representation of 3D micropatterned substrates capable of providing 3D geometric cues to DCs. DCs cultured on substrate with 3D geometric cues show podosomes aligning along the edges of the 3D micropatterns, indicating spatial arrangement of podosomes by the geometric cues. Cells were fixed and stained with phalloidin-Texas Red and an anti-vinculin mAb to visualize actin (red) and vinculin (green), respectively. The dotted lines in the insets represent the position of the edges of the micropatterned substrate.90 (D) DCs stimulated with PGE2 for 5 min and 16 h led to loss of podosomes and appearance of focal adhesions. Here, iDCs were seeded on fibronectin-coated coverslips and left untreated or stimulated with PGE2 then stained with an anti-vinculin mAb (green) and phalloidin-Texas Red (to detect F-actin, red).91 (E) Cluster disruption (CD) of DC led to maturation of DCs with exhibition of morphological changes as induced by LPS. DCs matured by DC or LPS were labeled for MHC II(red) and lysosomal marker Lamp2 (green).92 (F) SEM micrographs of spherical microparticles (top left), non-spherical 3X stretched microparticles (top right), spherical nanoparticles (bottom left), and non-spherical 3X stretched nanoparticles (bottom right). |
Despite the success and wealth of knowledge that has been gained using 2D or micropatterned 2.5D in vitro culture systems, they do not accurately represent structure of tissues in vivo, driving the emergence of works that use various 3D culture models which are important for understanding how change in mechanical properties of tissues in vivo affect cellular function.105,106 In comparison to culture on 2D, murine bone marrow derived iDCs cultured on 3D collagen scaffolds expressed similar levels of CD11b and costimulatory molecules but lower CD11c and MHC-II.60 When the iDCs matured, it was observed that expression levels of MHC-II, CD40, and CD83, although noticeably markedly upregulated on 2D, were only slightly upregulated when cultured in 3D, indicating the 3D collagen scaffold significantly altered the activated DC phenotype.60 Fang et al. also found that there was large variation in gene expression profiles between iDCs cultured on 3D collagen scaffolds vs 2D tissue culture dishes, with iDC related genes such as Ccl3, Ccl4, Ccr1, Ccr3, and Ccr5 being significantly upregulated when cultured on the 3D scaffolds.60 mDCs cultured on 3D collagen scaffolds also secreted higher IL-10 and lower levels of IL-12p70, IL-2, IFN-γ and TNF-α than those cultured on 2D.60 We have similarly shown that the cell culture dimensionality, and matrix density differentially affect DC differentiation and maturation. From tissue-centric perspective, expression of maturation surface markers (CCR7,CD209,CD80, CD86 and MHC-II) and secretion of cytokines for both iDC and mDC was found to be higher on 2D and differentially regulated by varying matrix density in 3D, with mDCs having increased CD80 expression and reduced CD86, CD11c and MHC-II expression in dense matrices.62,107 In contrast to what was observed with the other DC surface markers, CD11c was found to be more expressed in 3D matrices than on 2D.62 When pre-differentiated on 2D tissue culture plates and then transferred to varying substrates, the cell-centric perspective, both iDCs and mDCs showed enhanced expression of all surface markers and showed distinct cytokine secretion profiles when transferred from 2D into the 3D matrices.62 CD80 was expressed higher in iDCs in 3D, when compared to mDCs at the same conditions, while iDCs expressed CCR7 at a similar level to that of mDCs.62 These 3D studies, overall, stress on the importance of tissue dimensionality and stiffness in regulating DC behavior.
The impaired immunity faced by astronauts is akin to the poorer immune potency of aged individuals and perhaps also similar to patients with cancer. DCs, when exposed to simulated microgravity, have altered transcriptome profiles and were found to have reduced secretion of proinflammatory cytokines.70 In iDCs, simulated microgravity reduced the expression of CD11c, CCR7, and CD86 while CD209, MHC-II, and CD80 expressions were not affected.70 Similarly in mDC, simulated microgravity reduced expression CD11c, increased CD80 expression, but did not affect CD209, MHC-II, and CD86 expression.70 It has been reported that the impairment in maturation of BMDC upon exposure to simulated microgravity is mediated by an alteration in the NFκB signaling pathway.108
For most part, in tumors, DC maturation is induced by uptake of dying tumor cells.109 However, it has been reported that DCs also acquire antigens through direct DC-tumor cell or DC-DC interaction via a process known as nibbling, where there is transfer of plasma membrane and intracellular proteins from tumor cells or mature DCs to immature DCs.109,110,111 There are no reported studies to date that indicate if mechanical forces during these interactions dictate downstream DC biology. Although pathogen recognition and processing is the main inducer of DC maturation, there have been observations of DC maturation in the absence of inflammatory or microbial stimuli.92,112 This form of DC maturation is independent of TLR signaling. Instead, it is a result of the disruption of DC-DC adhesion mediated by E-cadherin, leading to the activation of the β-catenin pathway.92 These mDCs were shown to possess typical features of DC maturation including upregulation of MHC, costimulatory molecules (Fig. 3E), and chemokine receptors. However, they exhibited an entirely different transcriptional profile, failed to release inflammatory cytokines and elicited regulatory T cell responses rather than effector, indicating that they are involved in immune tolerance.
2.3. Effects of substrate dimensionality and stiffness on DC antigen uptake
Antigen uptake is another main function of iDCs and occurs through various mechanisms including phagocytosis. Through this mechanism, DCs engulf particulate antigens, by receptor-mediated endocytosis where DCs internalize soluble antigens that bind to receptors such as Fcγ receptors (FcγRs) and CLRs. Another mechanism is macropinocytosis, in which DCs take up soluble antigens by non-specifically internalizing large volumes of extracellular fluid.113,114,115 The protrusive dendritic structures formed by DC podosomes facilitate antigen sampling from areas deeper within the substrate by locating the spots of the least physical resistance (Fig. 4A).48 Formation of these protrusive structures also causes a localization of pattern recognition receptors, triggering localized antigen uptake, processing and presentation to T cells (Fig. 4B).48 The ability of iDCs to take up antigen is also significantly affected by its mechanical microenvironment, indicating possible mechanisms by which DC functions could be altered in the TME.
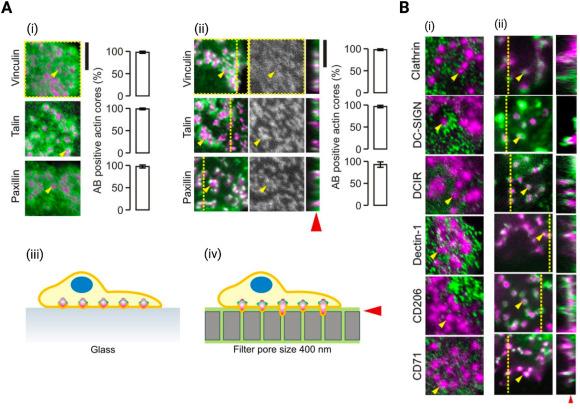
Fig. 4. Podosomes locate spots of least physical resistance and contain pattern recognition receptors to facilitate antigen sampling |
When cultured on PAA substrates of stiffness 12 kPa, DCs have been shown to significantly reduce expression of CLRs, CD209 and CD206, compared to being cultured on 2 and 50 kPa substrates.99 In line with the decreased expression of CD206 and CD209, antigen uptake was also significantly reduced by conditioning using 12 kPa PAA, suggesting a direct correlation between the expression of these CLRs and antigen uptake dependent on the substrate stiffness.99 On 3D collagen scaffolds, murine iDCs showed lower antigen uptake capacity compared to those cultured on 2D.60 van den Dries et al. also cultured iDCs on 2D micropatterns which provided geometric cues, promoting podosome formation in iDCs (Fig. 3B(ii)). They found that culture on the micropatterns triggered higher internalization of bacteria, indicating the important role of substrate geometry and podosome formation during antigen uptake.90 We have previously reported that for DCs derived and differentiated from monocytic cell lines in respective substrates, iDCs on 2D culture plates exhibit higher antigen uptake than in 3D matrices.62 However, in 3D microenvironment, antigen uptake was found to decrease in denser matrices compared to loose matrices. Given the heightened matrix density observed in the TME, these findings underscore the potential role of increased matrix density in the TME in compromising the functions of DCs. Additionally, when differentiated on tissue culture plates and transferred to respective substrates, antigen uptake by iDCs is reduced when transferred into new microenvironments.62 Upon immobilizing 3D collagen matrices with different molecular weights of HA, we observed that matrix bound LMW HA enhanced the antigen uptake capacity of iDCs via upregulation of the CD206 receptor.67 Finally, under simulated microgravity culture, we found that antigen uptake by iDCs was not significantly affected.70
Aside from mechanical properties of the microenvironment, properties of antigens such its geometry and size also affects antigen uptake by iDCs and immunomodulates as well. By comparing levels of phagocytosis of nanoparticles vs microparticles in combination with various adjuvants and differently shaped vaccine delivery vehicles (Fig. 3F), Matheas et al. showed that spherical nanoparticles combined with high molecular weight polyinosine-polycytidylic acid adjuvants were more efficiently internalized by immortalized murine iDCs (JAWS II mouse immature DC line) with induction of higher expression CD83 and CD86 maturation markers compared to elongated nanoparticles or microparticles with the different adjuvant combinations.98,116
2.4. Migration dynamics of DCs as influenced by shear forces and substrate factors
Migration is also an important part of DC function, whether migrating through the TME to capture tumor antigens, or migrating to lymphoid tissues following its maturation to activate the adaptive immune response. Rho-family GTPases are known to play a central role in DC migration: specifically, Rho activation causes DC extensions to retract while Rac activation causes the extensions to protrude further.117 Following their uptake, processing of antigens, and maturation, DCs become highly migratory and enter the lymphatic vessels through which they migrate to the lymph nodes where they activate T cells.118,119 Their migration to and through lymphatic vessels is known to be guided by chemokine signaling involving CCR7 receptors and chemokines such as CCL19 and CCL21.120,121,122 Substrate stiffness has been shown to influence the expression of CCR7 thereby altering their guided migration by CCL21 signaling, suggesting that tissue stiffness may influence the ability of mDCs to reach the lymphatic vessels.99 mDC migration to the lymphatic vessels is also facilitated by their initial docking to the lymphatic endothelium receptor LYVE-1 via its ligand hyaluronan which are binded to CD44 on the surface of mDCs.123,124 mDCs enter the lymphatic vessels through specialized button-like flap valves of about 2-3 μm in diameter.125 Increasing evidence also suggests that the flow of interstitial fluid towards the lymphatic vessels may affect the signaling induced by chemokines and ultimately plays a role in guiding DCs towards the lymphatic vessels, perhaps through a mechanism term autologous chemotaxis.126,127,128 Autologous chemotaxis is a process, whereby cells secrete chemokines and interstitial fluid drainage towards lymphatics generates a local chemokine gradient through which cells perform directed migration towards lymphatics.129 This has been shown in MDA-MB-231 breast cancer cell line, which possesses CCR7 and autologously secretes CCL19, and exhibited directed migration in response to flow (Fig. 5A).93 To mimic the intensity of shear stress generated in inflamed tissues such as tumors, Kang et al. developed a microfluidic channel with which they generated shear stress with intensity within 0.2-0.6 dyne/cm2, similar to that in inflamed tissues (Fig. 5B).85 Using this microfluidic model, they found that shear stress did not alter the speed of migrating DCs but influenced their direction of migration,85 possibly through the aforementioned autologous chemotaxis. Another factor that influences the DC migration is their binding capabilities to the ECM and to other cells such as endothelial cells which are found in the lining of blood and lymphatic vessels. Compared to monocytes and iDCs, mDCs have been found to possess lower binding capability to vascular endothelial cells which is in part due to their downregulation of CD11a, CD106 (VCAM-1) and CD54 (ICAM-1)130,131 which are required for adhesion. Additionally, the membrane of mDCs possess relatively excess negative charges which causes them to experience powerful repulsive forces from negatively charged endothelial cells.131 Overall, the decreased binding of mDCs to vascular endothelial cells may facilitate their migration through lymphatic vessels. Furthermore, the pumping activity that causes flow of lymph may also lead to either increase or decrease in rate of DC transit depending on the velocity of lymph flow.128 Moreover, transmural flow has been shown to trigger lymphatic endothelium to increase expression of DC chemoattractant molecule CCL21 and adhesion molecule ICAM-1 and decrease expression of lymphatic junctional adhesion molecules PECAM-1 and vascular endothelial (VE)-cadherin (Fig. 5C).94 Combined, these events increase lymphatic flow, thereby facilitating DC transmigration into lymphatic vessels and transit across lymphatic endothelium.94 Upon reaching the lymph node, mDCs travel along the fibroblastic reticular cells (FRC) to reach T cell zones where they can locate and prime T cell response.132 Their migration along the FRC network is dependent on the interaction of the C-type lectin receptor, CLEC-2 which is expressed by DCs with podoplanin (PDPN) expressed on FRCs.133 The activation of CLEC-2 by PDPN triggers DC spreading through the downregulation of Rho activity and myosin light-chain phosphorylation and causes the formation of F-actin-rich protrusions via Vav signaling and Rac1 activation.133 Ultimately, this leads to a rearrangement of the actin cytoskeleton of DCs and promotes their migration along the FRC network.133
Migration of DCs in vitro is differentially affected by the substrate or 3D microenvironment within which they are cultured. In a study where DC migration on PDMS-coated coverslips vs micropost array detectors were compared, it was found that migration is influenced by geometry of ligand presentation (Fig. 5D).95 It was observed that reorganization of the actin cytoskeleton, myosin contractility and α5β1 integrin engagement played a significant role in DC's sensing of ligand geometry.95 In 3D microenvironments, DC migration occurs solely by the force of actin-network expansion instead of by integrin-mediated adhesion as observed on 2D substrates.134 Myosin-II dependent contractions are involved in transmigration of DCs through narrow gaps134 and have been shown to have an effect on cell shape, and migration speed.96 These myosin-II dependent contractions result in the formation of protrusive podosomes at the leading edge of migrating DCs, facilitating their motility.135 Perinuclear Arp2/3 also causes polymerization of actin around the nucleus, providing a force that compresses and deforms the nucleus in order to allow the cells to pass (Fig. 5E).96,136,137 During migration, compression of cell body or indentation of the plasma membrane of DC causes assembly of Wiskott-Aldrich syndrome protein (WASp) into dot-like structures, providing activation platforms for Arp2/3 nucleated actin patches which provides the force to create space and facilitate forward movement.138 Both podosome formation and nuclei deformation coupled with localized degradation of ECM using metalloproteinase MMP-14 ultimately result in efficient transmigration of DCs through narrow constrictions.48,135 Mechanical stiffness of the narrow contractions has also been shown to modulate DC migration. When exposed to a custom-designed gel confiners (Fig. 5F) with lower stiffness (1.2 kPa), murine iDCs exhibited higher mean track speeds of 6.1 ± 0.3 μm/min than those migrating through stiffer confinement (3.5 ± 0.1 and 2.8 ± 0.1 μm/min for gels with E = 8.0 and 18.1 kPa, respectively).65 Additionally, persistence time of iDC motility, a temporal measurement for propensity of migration in a particular direction, was increased under lower mechanical load compared to high mechanical load.65 From this, Choi et al. suggest that increased motility and persistent time under low mechanical loading, together results in an increased travel range for DCs to scan a broader area while the reduction in motility under higher mechanical loading causes DCs to perform more through scanning of a small area.65
2.5. Antigen presentation and T cell activation
Antigen presentation and activation of T cells by DCs is the most critical step in initiating the adaptive immune response. During antigen presentation, the antigen peptide-MHC complex and stimulatory molecules (such as CD80 and CD86) interact with T cell receptor (TCR) and costimulatory receptors (such as CD28) respectively.139,140,141,142,143,144 Given the discussion so far about how different mechanical microenvironments can affect expression of MHC proteins and costimulatory molecules, it is plausible to infer that different mechanical microenvironments may also differentially impact antigen presentation and T cell activation by DCs. Unfortunately, only a few reported works have studied the effect of microenvironment mechanical influence on DC antigen presentation.
Using a mixed lymphocyte reaction assay, it was found that mDCs cultured on 2D more effectively stimulated allogeneic CD4+ T cell proliferation compared to those cultured on 3D collagen scaffolds.60 This is consistent with earlier discussed results from this study that showed that mDCs cultured on the 3D collagen scaffolds secreted lower levels of IL-2 and IFN-γ and higher levels of IL-10 which is known to suppress T cell proliferation.60,145 From a tissue-centric perspective, we similarly found that mDC mediated activation and proliferation of T cells was significantly reduced in 3D matrices compared to 2D with no significant matrix density dependent difference in T cell proliferation.62 In the cell-centric perspective, mDCs appeared to lose their T cell activation capability when transferred into new microenvironments and induced T cells to secrete IL-17, rather than IFN-γ.62 These results are indicative of T cells differentiating towards Th1 and Th17 when co-cultured with mDCs from tissue-centric and cell-centric perspective, respectively.62
β2 integrins have been shown to restrict DC-mediated priming of T cells and their loss leads to an accumulation of DCs and an increased Th1 immune response.100 Varga et al. have also shown that their pharmacological activation results in a Macrophage antigen-1 (CD11b/CD18) mediated reduction in their T cell activation capacity.146 Although substrate stiffness regulates the expression of β2 integrins, interestingly human mDCs cultured on 2, 12 and 50 kPa PAA substrates did not show any stiffness dependent differences in ability to stimulate T cells.99 On stiff PDMS (50 kPa), however, murine BMDC exhibited enhanced antigen presentation capacity and T cell activation.104 Finally, Piezo1 has been found to be important in directing reciprocal differentiation of Th1 and Treg cells.147 Using a mouse cancer model, Wang et al. found that genetic deletion of piezo1 in DCs inhibited DC-mediated priming of Th1 cells but rather triggered increased differentiation of regulatory T cell subsets, thereby promoting tumor growth.147 These piezo1 deficient DCs secreted increased amounts of TGFβ1 but reduced IL-12, suggesting that the observed regulation of Th1 and Treg phenotype occurs through the polarizing cytokines TGFβ1 and IL-12.147
3. Dendritic cells as providers of mechanical forces
Aside from their sensitivity to mechanical forces they encounter, DCs possess biomechanical properties which enables them to act as force providers, providing biomechanical cues to their microenvironment, to other DCs, as well as other cells they interact with, particularly T and B lymphocytes. The biomechanical properties which DCs possess during their different differentiation stages are important factors facilitating the roles they play both in the innate and adaptive immune response. Hence disruption of these properties could lead to alterations in immune functions of DCs. Within the tumor microenvironment, vascular endothelial growth factor (VEGF) produced by tumor cells negatively impacts biomechanical properties of mDCs via an abnormal remodeling of F-actin cytoskeleton.148 F-actin activity has been shown to have direct correlation with DC deformability.149 Changes in biomechanical properties induced by VEGF include decreased electrophoretic mobility, increased osmotic fragility, decreased viscoelasticity/deformability, and decreased transmigration.148 These changes, altogether, contribute to the mechanisms by which tumors escape the immune system.148 Xu et al. also observed changes in biomechanical properties of DCs including decreased deformability and reorganization of cytoskeletal F-actin when cultured with K562 cells, a human chronic myeloid leukemia cell line.150 They further showed that culture with K562 cells led to alterations in cytoskeleton-related genes including cofilin1 and profilin1 and matrix related genes and proteins including TIMP1 and MMP9, providing a molecular basis for the changes in biomechanical characteristics of the DCs within the tumor microenvironment.150
Under normal conditions, mDCs are significantly stiffer, possess higher membrane fluidity and have reduced nucleus mobility compared to iDCs.144,149,151,152,153 During maturation, cortical stiffness of DC is increased via actin polymerization, in a substrate dependent manner.154 Various methods have been developed for evaluating biomechanical properties of cells including DCs, some of which are summarized in Fig. 6A. Many of the cell surface receptors that are involved with lymphocyte activation are mechanosensitive and hence biomechanical cues from antigen presenting cells, particularly DCs play crucial roles in regulating communication with lymphocytes during activation.155 For example, the increased stiffness of mDCs has been found to lower the threshold for activation of CD4+ T cells but not CD8+.154,156 The reorganization of cytoskeletal actin that occurs to cause stiffness increase in mDCs also leads to restraining of ICAM-1 lateral motility, thereby promoting ICAM-1 affinity to LFA-1 on T cells and hence the formation of stable mature immunological synapse (Fig. 6C and D).157,158,159,160,161,162,163. This formation of a stable immunological synapse enhances T cell proliferation. Optimal T cell activation also requires inactivation of LFA-1 on DCs, allowing DCs to actively control antigen-driven T cell proliferation.164 In addition, mDCs’ increased membrane fluidity facilitates their mobility as well as their ability to expose costimulatory molecules for recognition by T cells.152 Costimulatory molecules, CD80 and CD86, have also been shown to contribute to the mechanical strengthening of DC-T cell interactions, thereby amplifying T cell functional activation.165 Similar to their physical interactions with T cells, DC stiffness is also important for antigen presentation for activation of B cells.166
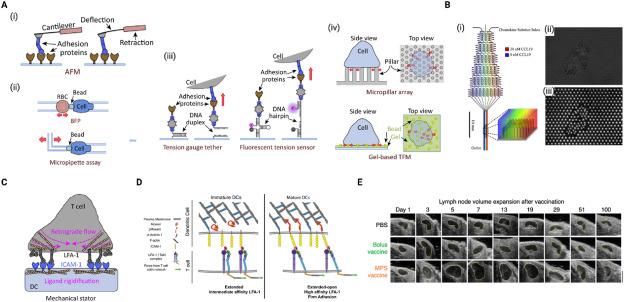
Fig. 6. Understanding biophysical properties and force providing capability of DCs |
In the lymph node, DCs exert tension on the FRC network, thereby contributing to lymph node expansion.169 Lymph node expansion is a characteristic of an ongoing adaptive immune response (Fig. 6E)170,168 and ultimately makes room for immune cells influx to the lymph node as well as for T cell proliferation within nodes. CLEC-2 on the membrane surface of DCs modulates stretching of the FRC network through interaction with PDPN on FRCs.169 CLEC-2-PDPN interaction triggers clustering and rapid uncoupling of PDPN from RhoA/C activation, thereby relaxing the actomyosin cytoskeleton and allowing FRC stretching.169
Force generation by DCs is also important during their migration.171 As discussed earlier, Arp2/3 complex mediated actin polymerization is essential for this force generation that facilitates DC migration.96,136,137,172 Using micropost array detectors incorporated into microfluidic gradient generators (Fig. 6B), Ricart et al. determined the traction forces generated by murine DCs during chemotaxis and chemokinesis to be 8 ± 1.4 and 16 ± 1.3 nN/cell.167 They show that migration of murine DCs is driven by short-lived traction stress (with a characteristic duration of 3min) concentrated at the leading edge which are dependent on actomyosin interaction rather than by chemical gradient.167,173 They also found that the maximum principal stress in the DCs occurred in front of the cell centroid and that using the spatiotemporal pattern of the traction stress, direction of future DC motion can be predicted.167
4. Lessons from DC mechanobiology: implications for the future direction of cancer therapy development
In the ever-evolving field of cancer research, DC mechanobiology holds profound insights into how DC functions may be impaired within the TME. Our understanding of the dynamic interplay between DCs and their mechanical environment comes with valuable lessons that could enable us to explore new targets and techniques for development of therapeutic interventions for diseases such as cancer which are characterized by increases in various forms of biomechanical cues. Generally, there are two main approaches that have been used to explore the engineering of DCs for cancer immunotherapy. The first involves direct in vivo targeting of intratumoral DCs in order to improve their immune function and enhance tumor control. For example, many studies have explored the direct in vivo targeting of DC cell surface receptors using ligands, adjuvants, or antibodies for either triggering proinflammatory cytokine secretion, blocking DC inhibitory signals or efficiently delivering tumor antigens to DCs, thereby improving tumor immune response.174,175,176,177 Others have also explored implantation of biomaterial-based scaffolds with the ability to release DC chemotactic agents or deliver tumor antigens for recruitment and maturation of DCs respectively.178 Despite substantial efforts devoted to the development of these strategies, the mechanobiology of DCs appears to have been overlooked in development of DC based therapies. We believe that with our understanding of DC mechanobiology, future therapeutic developments ought to explore the targeting of ECM properties as well as other biomechanical forces to modulate DCs in favor of tumor control. Theoretically, the mechanosensing machinery/pathways involved in DC response to TME biomechanical forces could be therapeutically manipulated to improve intratumoral DC functions and promote tumor control. Design of biomaterial based scaffolds for in vivo DC manipulation also need to take into account how the mechanical and topographical properties of the biomaterials can be leveraged to improve efficacy.179
The other approach which has been explored for dendritic cell-based immunotherapy is adoptive transfer of ex vivo activated and engineered DCs. This approach involves either isolating and expanding circulating DCs or differentiating DCs ex vivo from monocyte or hematopoietic precursors.180 The generated DCs are then manipulated, loaded with specific tumor antigens to induce maturation and are transferred back into the patient to induce antitumor immune response.181,182 As we showed that the inflammatory response for DCs transferred after differentiation and activation on tissue culture plates is attenuated, this mechano-immune memory advocates for cancer medicine to adopt lessons from DC mechanobiology. A recent study by Dombroski et al. proposed the use of fluid shear stress for cost effective activation of DCs for adoptive DC transfer therapy.183 Dombroski et al. showed that DCs exposed to sustained fluid shear stress using a cone-and-plate flow device exhibited immunophenotype characteristic increased activation.183 This work further brings to light the need to think about the mechanobiology of DCs in generation of DCs ex vivo and even in the design of carriers for adoptive transfer of the generated DCs. Our understanding of how geometric properties of antigens can affect DCs uptake, as discussed, also underscores the importance of a move towards design of well-defined antigen carriers for efficient loading of antigens onto DCs. Additional research is warranted to investigate the potential and leveraging various biomechanical cues for augmenting ex vivo activation of dendritic cells (DCs). Furthermore, there is a need to explore genetic modification techniques for manipulating DC mechanosensing machinery, aiming to enhance DC functions and improve their efficacy in tumor treatments.
Understanding DC mechanobiology can also enhance adoptive T cell transfer (ACT). ACT harnesses the power of autologous or allogeneic T cells, the primary effectors of the immune system, reengineers them, and expands them to target, overwhelm, and eliminate cancer cells.184 The adoption of ACT has significantly improved the success rate for hematological tumors and multiple myeloma and is paving the way into the treatment of solid tumors.185,186 Manufacturing of the T cells begins with the isolation of T cells from the blood. These T cells are subsequently activated/expanded using artificial antigen-presenting cells (aAPC), or DC surrogates, and additional modifications are performed before the final step of transferring the modified T cells back to the patient.187 aAPCs are essentially artificial cells that allow ex vivo mimicking of the cell-cell interaction that occurs between APCs and T cells, thereby providing stimulatory signals that trigger T cell activation and expansion.188,189 Clinically, magnetic beads known as dynabeads conjugated with anti-CD3 and anti-CD28, are used for the activation/expansion of T cells, thereby generating large numbers for transfer to patients.187,190 However, given the earlier discussion on the importance of the force providing capabilities of APCs such as DCs in activating T cells, it is clear that specific biomechanical properties of aAPCs are essential to achieve efficient ex vivo activation/expansion of T cells. Dynabeads, on the other hand, are made of high-stiffness materials191 and hence do not accurately mimic the biomechanical rigidity of native DCs.144,184 This idea has sparked numerous studies from us and others, investigating how the stiffness, dimensionality, shape and size of aAPC could impact T cell activation,189,192,193,194 thus shaping the criteria for the ideal aAPC design. We previously reviewed such studies extensively.144 Once more, this comprehension emphasizes the ongoing need for dedicated efforts to unravel the intricacies of DC mechanobiology, emphasizing its profound implications for the advancement of diverse forms of cancer immunotherapy.
5. Conclusions and future directions
In this review, we have extensively examined the diverse studies that have contributed to our present comprehension of how DCs are influenced by different biomechanical cues. Emphasizing these discussions, we acknowledge the distinctive biomechanical properties prevalent in the TME that are capable of modulating DC behavior. Thus, we underscore the significance of comprehending DC mechanobiology as a crucial element in unraveling how biomechanical cues within the TME may intricately shape/alter DC behaviors. Moreover, we delve into the implications of this knowledge, exploring its potential applications in therapeutic interventions for cancer, spanning from the direct targeting of endogenous DCs within tumors to influencing the manufacturing and delivery of DC vaccines, and finally, shaping the design of artificial antigen-presenting cells for adoptive T cell transfer therapy. Despite the considerable efforts in understanding DC mechanobiology, it is regrettable that our comprehension of this field lags behind that of other immune cells like macrophages and T cells, and therefore the therapeutic potentials within the DC realm remain largely unexplored. It is still not fully understood and characterized if specific mechanical cues could result in DC phenotypes representing specific DC subtypes discussed earlier. Additionally, it is unknown if the various DC subtypes will respond to various forms of mechanical stimuli in the same way. Our current understanding of DC mechanobiology also fails to account for the fact that DCs are not subjected to a mere single mechanical stimulus. Rather tumors and other tissues where DCs experience microenvironmental cues are made up of multiple biomechanical cues which may simultaneously impact DCs and integratively modulate their behavior. Perhaps, DCs may possess mechanisms whereby immunomodulation by different forms of biomechanical cues at a time may result in no effect at all or a different form of response altogether. These are very much unexplored questions. With the growing popularity and success of in vivo tumor models employing lysyl oxidase inhibitors to reverse tumor stiffness, it becomes intriguing to explore how this reversion of stiffness influences the functioning dendritic cell populations within the broader context of other tumor-associated factors, and how this may contribute to the improved responses to immunotherapy that is observed.195,196,197 Given the importance of DC mechanobiology studies and the therapeutic implications discussed, it is imperative for upcoming research endeavors to address these unanswered questions and propel the advancement of our understanding in this field. Finally, the clinical application of DCs for cancer therapy, as well as other cell-based therapy, has largely overlooked mechanobiology. Our hope is to advocate the need for translational medicine to incorporate mechanobiological considerations in the development of cell-based therapeutics.
Ethical approval
This study does not contain any studies with human or animal subjects performed by any of the authors.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgment
The authors acknowledge the support from New York University Abu Dhabi (NYUAD) Faculty Research. Fund (AD266). We further acknowledge NYUAD Office of Undergraduate Research for facilitating and the Office of Global Education for funding the Visiting Undergraduate Research Program.
Abbreviations
aAPC artificial antigen presenting cells
APC antigen presenting cells
Arp2/3 actin-related proteins-2/3
BMDC Bone marrow derived dendritic cells
CCL19 Chemokine ligand 19
CCL21 C-C motif chemokine ligand 21
CCR7 C-C chemokine receptor type 7
Ccr3 C-C chemokine receptor type 3
Ccr5 C-C Motif Chemokine Receptor 5
CD106 Cluster of differentiation 106
CD11a Cluster of differentiation 11a
CD11b Cluster of differentiation 11b
CD11c Cluster of differentiation 11c
CD18 Cluster of differentiation 18
D209 Cluster of differentiation 209
CD28 Cluster of differentiation 28
CD40 Cluster of differentiation 40
CD54 Cluster of differentiation 54
CD80 Cluster of differentiation 80
CD83 Cluster of differentiation 83
CD86 Cluster of differentiation 86
CLR C-type lectin receptors
DC Dendritic cells
ECM Extracellular matrix
FRC Fibroblastic reticular cells
GTPases GTP-binding proteins
HA Hyaluronic acid
HMW High molecular weight
ICAM-1 Intercellular Adhesion Molecule 1
IFN-γ Interferon gamma
(IL)-1α Interleukin 1α
IL-12/IL-12p70 Interleukin 12
IL-17 Interleukin 17
IL-1β Interleukin 1β
IL-6 Interleukin 6
iDC Immature dendritic cells
LFA-1 Lymphocyte function-associated antigen 1
LMW Low molecular weight
LPS Lipopolysaccharide
LYVE-1 Lymphatic endothelium receptor
MHC Major histocompatibility complex
MMP-14 Matrix metalloproteinase-14
NLR nucleotide-binding oligomerization domain-like receptors
PAA Polyacrylamide
PDMS Polydimethylsiloxane
PDPN Podoplanin
PECAM-1 Platelet endothelial cell adhesion molecule
PGE2 Prostaglandin
Rac1 Ras-related C3 botulinum toxin substrate 1
RhoA Ras homolog family member A
RPM Random positioning machine
SEM Scanning electron microscopy
TCR T cell receptor
Th17 T helper 17
Th1 T helper 1
TIMP1 Tissue inhibitor of metalloproteinases 1
TLR Toll-like receptors
TME Tumor microenvironment
TNF-α Tumor necrosis factor alpha
VCAM-1 Vascular cell adhesion molecule 1
VEGF Vascular endothelial growth factor
WASp Wiskott-Aldrich syndrome protein
YAP Yes-associated protein

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}