
1. Introduction
Cells respond to mechanical stimuli (e.g., changes in substrate stiffness) by altering their mechanical state (e.g., cell stiffness, motility, adhesion, and contractility).1 These phenotypic properties of cells are modulated by the organization of the cytoskeleton,2 which plays a significant role in mechanosensing.3,4,5 Dynamic reorganization of the cytoskeleton is a part of cellular homeostasis and often goes awry in cancer. Deregulated cytoskeletal assembly is implicated in tumor cell migration, invasion, and metastasis.6,7,8
Communication between a cell and its surroundings is potentiated by trafficking that involves membrane environments. Endocytosis causes the internalization of cargo at the plasma membrane and contributes to several cellular processes.9,10 Endosomal cargo is recycled to the plasma membrane or slated for lysosomal or secreted extracellularly in the form of exosomes. Endocytic pathways are deregulated in cancer cells:11 enhanced integrin recycling promotes invasive migration,12,13 mechanisms that undermine lysosomal degradation of growth factor receptors (such as EGFR and MET) enhance proliferation,14 altered E-cadherin trafficking induces epithelial-to-mesenchymal transition,15,16 and differential expression of chemokine receptors are observed during metastasis.17,18,19
Among various trafficking vesicles in the extracellular microenvironment, exosomes are 100 nm vesicles generated through the endocytic pathway and secreted via exocytosis.20,21 Exosomes are efficient potentiators of cell-cell communication, with the exosomal cargo actively mediating signaling between parent cells and recipient cells.20 Tumor exosomes are implicated in cancer progression and metastasis, their roles in tumor cell invasion,22 migration,23,24 and immune interactions25,26,27 are being appreciated with recent and ongoing studies.
From endocytosis at the plasma membrane to exosome generation at the endosomes, membrane curvature remodeling is an essential phenomenon in trafficking pathways. Among several mechanisms for membrane remodeling,28 peripheral membrane proteins capable of generating curvature play a central role: these include epsin N-terminal homology (ENTH) domain 29, Bin-Amphiphysin-Rvs (BAR) domain 30, Endosomal Sorting Complex Required for Transport (ESCRT) III protein complexes,31 to name a few. Curvature generation by these proteins will depend on the membrane's mechanical properties (such as tension and bending rigidity). Moreover, forces from the cytoskeleton will influence membrane deformability.32,33 Cytoskeleton-related tension will, therefore, affect the function of curvature-inducing proteins.
Altered ECM stiffness in tumorigenesis will influence cytoskeletal dynamics to define a new mechanosensitive mechanism in tumor cells for regulating trafficking pathways. Specifically, this mechanism can connect cortical tension and vesicle biogenesis, conferring an intriguing mechanosensitivity paradigm in tumor initiation and progression. Intracellular transport mechanisms (endocytosis, exocytosis, endosomal sorting, and exosome release) dynamically regulate and impact signaling by modulating receptor activity and protein localization.34,35 Our outlook is to bridge molecular and cellular principles surrounding membrane curvature generation with rewiring of cellular signals. Here, we describe a general framework that integrates signaling with other cellular functions like trafficking, cell-cell and cell-matrix adhesion, and motility. We are guided by how a physical change in cell membrane configuration caused by external stimuli alters trafficking and signaling. We apply this framework to explore the link between trafficking, signaling in the tumor microenvironment, and cell fate.
2. Multiscale computational approach
We adopt a multiscale computational approach to enable the bridging between molecular mechanisms of curvature sensing, cellular localization, and curvature generation and the subsequent link to trafficking and signaling. Coarse-grained molecular dynamics simulations were performed in order to determine the molecular mechanisms of curvature generation and sensing. Established simulation protocols (shared at github.com/bradleyrp/automacs) make it possible to simulate protein/lipid systems at many concentrations systematically. The resulting MARTINI/GROMACS simulations scale linearly on high-performance computing platforms, making generating these simulation data efficiently possible. To predict the deformation fields for a particular protein-bilayer system, we simulated it using coarse-grained simulation methods. Specifically, we conducted the CGMD simulations of the protein domains embedded in mixed DOPC: DOPS bilayers with physiological concentrations of phosphoinositide lipids (1-4%)36,37,38 using the MARTINI coarse-graining schemes and force fields.39,40,41 Each protein is bound to a single PI(4,5)P2 molecule parameterized from atomistic simulations which remains bound to the amphipathic helix H0 near the known binding pocket.29 Simulations lacking PI(4,5)P2 failed to show measurable curvature and in some cases the ENTH domains even detached from the bilayer. This result is consistent with experiments that show that ENTH domains require binding42 and motivates our use of a single PI(4,5)P2 which is sufficient to retain ENTH at the bilayer in our model. The sensitivity of our results to the optimization parameters were tested extensively and the robustness of the results was confirmed in our prior work.43
We require a similarly systematic method for measuring the deformation field to complete the molecular portion of the simulation-curvature-morphology calculation. We adopt an atom-to-field mapping, which translates the structural details of the membrane at the molecular level to phenomenological parameters at the mesoscale and describes a methodology that can investigate the membrane protein system at both these scales and transfer information between them. We have previously estimated curvature fields induced in protein-bilayer simulations for ENTH domains44 and the protein Exo 7045 by directly analyzing the statistics of deformations in the simulation trajectories. For our protein-bilayer system consisting of 1 million particles, a typical dynamics run for a length of 500 ns required 48 h of computing time on a 48-core (3 nodes, 16 processors each), therefore requiring 1920 CPU hours. In principle, the methods have linear scaling for 1000s of cores enabling exploration of large systems.46
Our studies also revealed that the induced curvature is of the same scale as natural undulations, making it challenging to convolve the two effects.47 To overcome this challenge, we utilize an analysis method for a molecular description of protein-induced bilayer curvature where we quantify the protein-induced deformation at low surface coverages in a CGMD simulation using a continuum model based on the Canham-Helfrich Hamiltonian, which relates these deformation fields to height fluctuations in the bilayer in the presence of an intrinsic curvature field.48,49 The atom to field mapping and the different ways to compare to experiments are discussed in two earlier works.43,44 The main limitation is that single molecule studies, while most quantitative, may fail to capture the mapping because of cooperativity which is hard to decipher especially a priori. On the other hand, cellular experiments which include the cooperativity may not yield a quantitative estimate of curvature at the nanoscale.
At the micron scale, where a cell and its internal organelles display extraordinary complexity in their shape and organization, the physical properties of a biological membrane were explored using phenomenological continuum scale models. We have a generalized approach to studying membrane morphologies under high curvature such that they can be successfully applied to morphological transitions in cell membranes (such as vesiculation and tubulation).44,49,50 Using the machinery of dynamically triangulated Monte Carlo or DTMC simulations,50,51 we explored the roles of cooperative protein-protein and protein-membrane interactions clathrin-mediated endocytosis.52 In our model, multiple epsins localized spatially and orientationally templated on the clathrin coat collectively play the role of a curvature-inducing coat.
We have also successfully extended these results to consider how thermal fluctuations influence the vesicle morphologies,44,52,53 see Fig. 1 (left). We have similarly applied the anisotropic curvature model in the DTMC simulations to capture the collective role of Exo 70 domains found in the Exocyst complex proteins in reshaping membranous vesicles. When attached to the outer surface of giant unilamellar vesicles (GUVs), wildtype proteins induce inward-growing tubes. This phenomenon is predicted in our simulations by choosing a negative value for the anisotropic curvature field,45 see Fig. 1 (middle); in particular, the equilibrium membrane conformation stabilized in the model shows striking similarity to that obtained from experiments. In experiments, it has been observed that the ESCRT proteins are localized to the neck region of the budded vesicle.54 Using the anisotropic curvature model and localizing the protein curvature field to an annular region, we demonstrated a novel phenomenon where a negatively curved vesicular bud is formed due to the constrictive action of either positively or negatively curving proteins, see Fig. 1 (right).
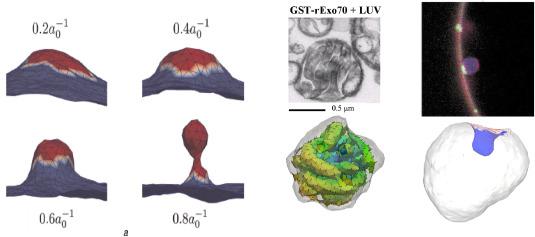
Fig. 1. (Left)membrane deformation profiles under the influence of imposed curvature of the epsin shell model for three different coat areas; here κ=20 kBT. For the largest coat area, the membrane shape is reminiscent of a clathrin-coated vesicle. Here, the vesicle morphologies subject to thermal fluctuations and entropic effects. (Middle) experimental observation of Exo 70 on GUV and (Right) results from the anisotropic curvature model. (Right) Fluorescence image shows the membrane in red and the location of ESCRT proteins in green; simulation results showing the novel phenomenon of budding by constriction – negative curved bud generated by negatively curved proteins localized to an annular region; the protein field does not localize the area of the bud. |
3. Results
3.1. Geometry through extracellular matrix (ECM) dimensionality tunes cytoskeletal tension to modulate cellular stress
Kai et al. investigated how engagement with a three-dimensional extracellular matrix influences cellular stress in cancer cells.55 They observed that cells within a three-dimensional extracellular matrix decreased and redistributed the actin crosslinker filamin to reduce their cortical tension. They studied the effect of cortical actin tension on membrane morphology using mesoscale models described in section 2. Cortical actin regulates membrane protrusions through membrane pinning, influencing membrane topology.56 Membrane topology controls the binding of curvature-generating proteins and cell signaling and the trafficking of cellular proteins, including their recycling and secretion.57 Consistent with these observations, cells engaging the ECM in 3D increased recycling and secretion (SEC61 expression), suggesting cortical actin tension regulates secretory protein translocation into the ER lumen. DTMC simulations were conducted to explore the morphology of highly curved membranes.44,49,50 Using this framework, Kai et al. quantified the surface deformations induced by membrane proteins as curvature fields and studied the emergent morphologies of the membrane using isotropic and anisotropic curvature models. They simulated membrane patches with different excess areas, which is the variable conjugate to the membrane tension and depends on cortical actin tension.58 The model predicted that the protrusions in cells engaging the ECM in 2D (reflecting high cortical actin tension) would be shorter and more transient. By contrast, the membrane protrusions in cells engaging the ECM in 3D (reflecting lower cortical actin tension) would be longer and more stable. Consistent with these predictions, live cell microscopy showed that the cells in 2D formed dynamic, short membrane protrusions, whereas cells in 3D formed longer, more stable protrusions.
Plasma membrane protrusions are cylindrically shaped. While the neck or annulus of the protrusion generates positive membrane curvature, the cytoplasmic length of the protrusion generates a negative curvature. Membrane curvature regulates the binding affinity of membrane curvature-associated proteins.28 To explore this relationship, the DTMC model was used to predict protein recruitment. Kai et al. computed the free energy to recruit a protein to these three spatial regions (protrusion, annulus, and basal) as a function of cortical actin tension. The calculations show a more favorable recruitment of such proteins in 3D than in 2D. Given that Exo 70 is a negative curvature-inducing protein that is also a component of the exocyst complex targeting secretory vesicles to the plasma membrane, Kai et al. predicted that Exo70 could be regulated by cortical actin tension to facilitate secretory protein trafficking.45 Consistent with model prediction, in the experiments of Kai et al., Exo70 showed better plasma membrane localization in 3D (low cortical tension) as revealed by a strong co-localization of the EGFP-Exo70 with membrane-tagged mCherry as compared to minimal membrane associations observed in 2D. The data suggest cortical tension regulates secretory protein trafficking to the plasma membrane by controlling plasma membrane protrusion activity and topology.
The results implied that how a cell ligates its ECM (2D or 3D) profoundly affects ER function and protein secretion. The results implicated filamin-dependent cortical actin tension as a critical regulator of this ECM ligation-mediated ER homeostasis. Prolonged ER stress responses compromise cell viability, and impeding secretory protein trafficking can severely compromises ER function.59 Cells with high cortical actin tension had significantly reduced cell viability compared to cells with low cortical actin tension. Accordingly, the viability of cells in 2D could be significantly extended if their myosin activity was inhibited. Also, knocking down the negative curvature-generating binding protein Exo 70, which has been implicated in protein secretion and membrane topology regulation, severely compromised the viability of the cells. The studies underscore the importance of geometrical context in organelle homeostasis and tissue behavior. Cells in 3D showed reduced endoplasmic reticulum stress response signaling, secreted and assembled a polarized endogenous basement membrane, and survived better under chemotherapeutic and radiation doses.
3.2. Regulation of trafficking vesicles by ECM stiffness
One of the main findings of the application described in section 3.1 is that trafficking vesicles potentiate the link between the dimensionality of ECM engagement and cell survival. Therefore, this section describes a causal mechanism that further illuminates this link.60 Cells respond to mechanical stimuli by altering their mechanical properties, such as cell stiffness, motility, adhesion, and contractility.1 The mechanical properties of cells are modulated by the organization of the cytoskeleton.2 From endocytosis at the plasma membrane to biogenesis of exosomes as intraluminal vesicles in late endosomes, membrane curvature remodeling is an essential component of trafficking pathways. As discussed in section 3, curvature generation by these proteins will depend on the membrane's mechanical properties (such as tension and bending rigidity). Forces from the cytoskeleton also strongly influence membrane deformability by modulating the effective tension in the membrane through membrane-to-cortex attachments.32,33 Cytoskeleton-related tension will, therefore, affect the curvature generating capabilities of the curvature-inducing proteins.
It has been postulated that cell-cell communication can be tuned through mechanisms of mechanosensitive biogenesis of trafficking vesicles.20,59 Exosomes are nano-sized vesicles produced through the endocytic pathway and secreted via exocytosis, are efficient mediators of cell-to-cell communication,20,21,61 and tumor-derived exosomes have been implicated in cancer progression,61 tumor cell invasion,22,23 metastasis,24 and evasion of immune surveillance,25,26,27 thereby linking to several cancer hallmarks. We thus wondered whether stiff ECM affects the membrane curvature generation needed in trafficking pathways through its effect on cortical tension.62 Parihar et al.,62 studied the dependence of cell mechanics of different cancer cells on ECM stiffness and composition to answer this question; this study reported extensive analysis on the physical behavior (namely, cell stiffness) spanning different cancer cell lines grown on different ECM stiffness and composition. The study utilized the biophysical modeling framework involving the DTMC-based mesoscale membrane model to elucidate how ECM mediated changes in cortical tension of cells can influence production of vesicles.
While another study reported the experimental results for the ECM dimensionality corresponding to a particular cell type,55 the mechanistic insights from the accompanying biophysical model (similar to the one mentioned above) about how the change in cytoskeleton tension due to ECM dimensionality effects membrane protrusion can be extended to other cell types.
In order to understand which proteins could be implicated in cell response to mechanosensing, the authors62 analyzed a proteomics dataset which involved the study of 20 normal and cancer cell lines in seven different ECM substrates. They focused on the list of cytoskeletal-related proteins in modulating cell stiffness in response to ECM mechanics, including the cytoskeleton's major components. A neural network with these protein counts as input predicted with balanced accuracy (BACC) of 98% on a test dataset whether a cell paired with a given substrate was likely to belong to the low cortical stiffness or high cortical stiffness category.62 This analysis reveals a highly diverse behavior among different cancer cell lines on varied ECM substrates, which can be captured through a low-dimensional fingerprint of 18 selected cytoskeleton-related proteins.
The study also conducted DTMC simulations to delineate how membrane deformations depend on cortical tension. Section 2 described the spatial localization of curvature-inducing proteins as a critical feature in the curvature generation mechanism. DTMC simulations have revealed a micellization-like mechanism for membrane tubulation, wherein tubes formed by self-assembling curvature-inducing proteins are analogous to micelle aggregates formed by self-assembling surfactant molecules.48 To address how the curvature-generating capabilities of curvature-inducing proteins are affected by cortical tension, DTMC simulations were used to explore the morphological conformational space of planar to highly curved membranes.44,48,50 The dependence of cortical tension is incorporated by simulating membrane patches subject to different excess areas, where increasing excess area corresponds to decreasing cortical tension.58 Parihar et al. analyzed how the membrane morphology changes with growing excess area for a given total number of protein-induced curvature fields. The theory they developed predicted a Hill-type ultrasensitivity for the effect of cortical tension on membrane vesiculation,62 and this behavior has since been verified experimentally.63
This tension-dependent recruitment of curvature-inducing proteins combined with our model's prediction of curvature generation suggests an interesting mechanosensitive axis that could be exploited to hijack intracellular trafficking mechanisms by cancer cells to gain fitness advantage. For instance, stiff ECM can increase the secretion of exosomes by cancer cells and also influence the composition of the bioactive cargo carried by exosomes.64 Specifically, exosomes produced by cancer cells on stiff ECM have been shown to increase proliferative signaling in the target cells,63 promote migration of recipient cells,65 and found to carry increased composition of specific adhesion receptors that mediate their organotropism for establishing pre-metastatic sites.66
To illustrate how essential signature proteins (Rab, SNX, EXOC and ATG) involved in regulating vesicle trafficking might be altered in the patients of cancer types, one can use the cBioPortal (http://cbioportal.org) to query for and analyze the mutations and copy number variations of the signature proteins in the extensive patient-derived cancer genomics data from The Cancer Genome Atlas (TCGA). Table 1 summarizes the results from this genomic analysis. We can see that a significant proportion of cancer patients across different cancer types showcase genotypic variations in the protein families critical in trafficking pathways, further highlighting the importance of the signaling (regulatory) portion of intra- and inter-cellular cargo trafficking machinery in tumorigenesis.
Table 1. Genomic analysis performed using cBioPortal on patient-derived cancer genomic data from TGCA PanCancer Atlas. Mut% and CNA% correspond to percentage of samples with one or more mutations and copy number alterations respectively. Samples profiled is the number of patient samples having the genomic data needed for that particular analysis. While max is the maximum value of Mut% (CNA%) for a protein family, ≥4% refers to the proteins having Mut% (CNA%) greater than 4%. Rab: Rab GTPases, SNX: sorting nexins, EXOC: exocyst complex components, and ATG: autophagy-related proteins. |
| Tissue Type | Samples Profiled | Empty Cell | Rab | SNX | EXOC | ATG | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| max | ≥4% | max | ≥4% | max | ≥4% | max | ≥4% | |||
| Skin | 440 | Mut% | 5 | Rab27B | 5.7 | SNX29, 13, 19, 31, 9 | 5 | EXOC1, 7, 4, 6B | 10 | ATG2A, 2B, ULK1, 2 |
| 367 | CNA% | 3.8 | 2.7 | 4.6 | EXOC3 | 3 | ||||
| Prostate | 494 | Mut% | 0.6 | 0.8 | 0.6 | 0.8 | ||||
| 489 | CNA% | 4.3 | Rab2A, 3C | 8.6 | SNX14, 31, 3, 16 | 2.2 | 8.6 | ATG5 | ||
| Ovary | 523 | Mut% | 0.6 | 1 | 1 | 0.8 | ||||
| 572 | CNA% | 10.8 | Rab8A, 19, 3D, 6A, 30, 25, 2A, 4A, 38, 3A, 13, 22A, 39B, 44 | 12.4 | SNX31, 27 | 9.1 | EXOC3, 4, 2, 8 | 7.9 | ATG4D, 9B, 16L2, 3 | |
| Lung | 566 | Mut% | 1.2 | 2.5 | 3 | 3.7 | ||||
| 511 | CNA% | 9.4 | Rab 13, 25, 22A, 29 | 10 | SNX27, 6, 16, 31 | 12.9 | EXOC3 | 2 | ||
| Colon | 534 | Mut% | 2.6 | 4.9 | SNX13 | 3.7 | 6.2 | ATGA2A, 2B, 9B | ||
| 592 | CNA% | 7.6 | Rab22A | 7.6 | SNX21 | 0.8 | 1.5 | |||
| Breast | 1066 | Mut% | 0.7 | 0.7 | 1 | 1 | ||||
| 1070 | CNA% | 9.2 | Rab4A, 29, 13, 25, 2A, 22A, 37, 26, 34 | 11.1 | SNX31, 27, 16 | 9.4 | EXOC8 | 4 | ATG16L2 | |
| Brain | 397 | Mut% | 0.5 | 1.3 | 1.3 | 2 | ||||
| 575 | CNA% | 1.4 | 1.7 | 5.2 | EXOC1 | 1.6 | ||||
4. Conclusion
A correlation exists between cancer cell mutations, which confer a fitness advantage to cells with a gain of essential cellular functions like cell adhesion and motility, signaling, transcriptional regulation, and intracellular trafficking. Defective signaling and vesicular trafficking of growth factor receptors and their subsequent alteration of cell fate by influencing the downstream signaling pathways are the hallmarks of cancer cells.14 The central goal of cancer biology is to find a mechanistic link between the above two factors, which has important implications for biomarker development and therapeutic efficacy. The main goal of oncology is to translate these findings into treatment regimens where optimization of therapeutic regimens can be customized to the genetic and physiological make-up of individualized patients or patient cohorts. Mechanistic studies that support the correlations with a causal link, such as those described here, can serve as a conduit for rational therapies.
Metastatic cancer is a major clinical challenge that accounts for over 500,000 deaths per year.67,68,69,70 Delineating mechanisms would identify actionable biomarkers and novel therapeutic targets. Tumors develop within a cellular microenvironment characterized by altered extracellular matrix stiffness and composition, blood vessel architecture, and immune cell interactions. These microenvironment components are causal in tumor amplification and metastasis.71,72,73,74 The immune cells that are initially recruited to the tumor are anti-tumorigenic; however, as malignancy proceeds, it shifts towards a pro-tumor phenotype. Also, the frequency of the tumor-infiltrating immune cells is reduced. The reprogramming of immune cells to a pro-tumor phenotype and immune cell depletion and exhaustion are traits associated with an aggressive metastatic tumor.75,76,77,78,79,80,81,82,83,84,85
A stiff extracellular matrix and cytoskeletal tension promote tumor cell growth and survival and stimulate cell invasion in culture.86,87,88,89,90,91,92,93 Studies in vivo using genetically engineered mouse models demonstrated that stiff ECM promotes malignant transformation.90,94,95,96,97,98,99 A definitive understanding of the interplay between protein binding/migration and membrane curvature evolution is emerging but still needs to be completed. The mechanisms underpinning such behavior are crucial in intracellular assembly and stability of organelles (which often sustain extreme curvatures), and in intracellular transport and sorting of proteins and cargo. Exosomes have drawn tremendous interest in cancer research as tumor-derived exosomes are implicated in modulating the tumor microenvironment, suppressing anti-tumor immunity, and preparing the metastatic niche for progression.26,27,100,101,102,103,104,105,106,107 We explored how tissue mechanics regulates cell cortical tension to influence exosome production and cargo composition by co-opting the biophysics and the signaling dynamics of intracellular trafficking pathways discussed in Section 3. How these exosomes contribute to suppressing anti-tumor immunity and promoting metastasis could be a subject of future investigations.
The impact of the described multiscale modeling approach goes beyond the fundamental investigation of specific cellular processes toward actual clinical trials. Exosomal mechanisms are critical to developing and approving liquid biopsy technologies, which are slated to be the future of non-invasive, longitudinal profiling of evolving tumors and resistance to therapies in cancer patients to bring us one step closer to the promise of personalized medicine.108 Moreover, delineating the mechanobiological regulation of exosomes is critical to our understanding of the evolution of solid tumors under physical stress from ECM and the interaction of tumors with the immune system, making it central for therapy, including immunotherapy. Already, clinical studies have revealed that exosomal profiling can classify responders and non-responders to immunotherapy in the treatment of some solid cancers.27
Ethical approval
This study does not contain any studies with human or animal subjects performed by any of the authors.
CRediT authorship contribution statement
Kshitiz Parihar: Conceptualization, Writing - original draft, Writing - review & editing. Seung-Hyun B. Ko: Writing - review & editing. Ryan P. Bradley: Writing - original draft. Phillip Taylor: Writing - original draft. N. Ramakrishnan: Writing - original draft. Tobias Baumgart: Writing - original draft. Wei Guo: Writing - original draft. Valerie M. Weaver: Writing - original draft, Writing - review & editing. Paul A. Janmey: Writing - original draft, Writing - review & editing. Ravi Radhakrishnan: Conceptualization, Funding acquisition, Writing - original draft, Writing - review & editing.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgments
We thank the Guo, Weaver, Radhakrishnan, and Janmey Labs members for insightful discussions related to the topics covered in this chapter. This study has received funding from the National Institutes of Health under R35GM136259 and U01CA250044. PAJ was also supported by the National Cancer Institute and Leidos Biomedical Research, Inc. under contract 15X008 with Frederick National Laboratory for Cancer Research. Computational resources were partly available from the extreme science and engineering discovery environment (XSEDE) under grant MCB200101.

{kind=link}
{kind=link}