
1. Introduction
G protein-coupled receptors (GPCRs) are the largest class of membrane proteins that function as major cell surface receptors in response to diverse extracellular stimuli, such as amines, peptides, and odor.1 The over 800 members human GPCRs can be structurally divided into five families: class A (rhodopsin-like), class B1 (secretin), class B2 (adhesion), class C (glutamate), and class F (frizzled) (Fig. 1), all of which contain a canonical seven-transmembrane (7TM) domain.2 For class A GPCRs, signaling is triggered by agonist binding to its orthosteric pocket, which results in rearrangements of the conserved 7TM to allow efficient coupling of the heterotrimeric G protein or β-arrestin;3,4,5,6 whereas for other families, signaling pathways are complexed, as their N-terminal extracellular region (ECR) also contributes to the recognition of their ligands.7 Meanwhile, allosteric ligands have been reported for all families, with their binding pockets varied around the 7TM of different receptors.
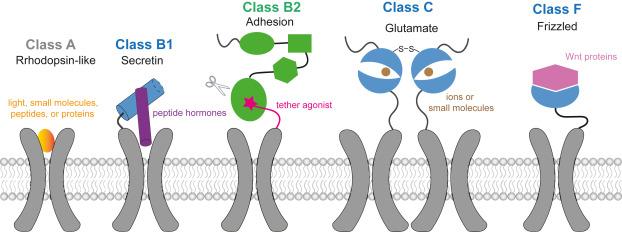
Fig. 1. Cartoon models for ligand recognition in different GPCR classes. |
Among non-class A GPCRs, adhesion GPCRs (aGPCRs) stand out because of their unique structure, featuring variable tandem adhesion domains, followed by a common GPCR autoproteolysis-inducing (GAIN) domain at their ECR (Fig. 2A).8 Primary, aGPCRs mostly interact with their ligands through their extracellular adhesion domains, although some endogenous ligands remain unidentified. Consequently, a major focus in the aGPCR field is the deorphanization of these receptors. Furthermore, several aGPCRs have been observed to respond to mechanical forces, either in the presence or absence of specific ligands, suggesting the complexity of the stimuli that trigger signaling in this receptor family.9,10,11,12,13 Recent studies have shown that aGPCRs play diverse roles, ranging from cell adhesion to immune defense and development, and consequently, their dysfunction is linked to various adverse health outcomes, such as inflammation, neurological disease, and cancer.14,15 Detailed examinations of the signaling pathways, functional consequences, and pharmacological properties of aGPCRs have been reviewed previously.16,17,18,19
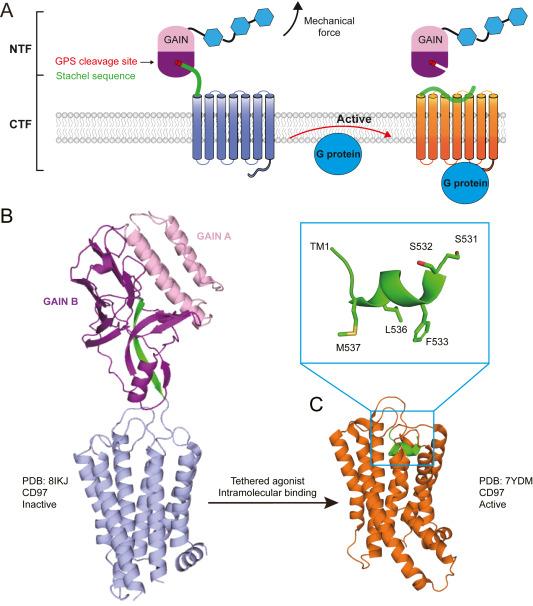
Fig. 2. Force-induced activation mode of adhesion GPCR. |
2. aGPCR and structures
Autoproteolysis occurs at a highly conserved motif in the GPCR proteolytic site (GPS) situated within the GAIN domain (Fig. 2). Structural analyses revealed that the GAIN domain can be divided into two subdomains: subdomain A, characterized by an α-helical structure and subdomain B, which predominantly comprises a β-sheet structure. Subdomain B notably contains the GPS motif, including the C-terminal sequence subsequent to autoproteolysis, termed the Stachel peptide, which folds within the β-sheet structure. Hence, despite autoproteolysis occurring in approximately 2/3 of aGPCRs during expression, the N-terminal fragment (NTF), encompassing the adhesion domains and the majority of the GAIN domain (excluding the Stachel peptide), remains associated with the C-terminal fragment (CTF), comprising the Stachel peptide along with the 7TM and intracellular region (ICR) on the cell surface (Fig. 2A). Ligand binding to the ECR was suggested to induce the release of the Stachel peptide, which then reorients and docks to 7TM, triggering downstream signaling pathways (Fig. 2A). Therefore, the Stachel peptide is also referred to as a tether agonist; previous studies have confirmed its agonistic function using peptides synthesized from several aGPCRs.20,21,22,23 This conformational change during activation is believed to be accomplished with the assistance of a mechanical force.13,17,24,25
With the application of lipid cubic phase (LCP) and T4-lysozome fusion technologies, the first engineered GPCR structure, β2 adrenergic receptor, was determined jointly by Kobilka and Stevens in 2007.26,27 Since then, hundreds of GPCR structures have been determined in complexes with different types of ligands by crystallography, and later by the cryo-EM method.28 In particular, since 2017, the cryo-EM method has been successfully used to determine the GPCR structure in complex with different downstream adaptors such as G proteins, β-arrestins, and GRKs. With these technological developments, the first transmembrane domain-containing adhesion GPCR structures were solved in complexes with different types of G proteins in 2022.29,30,31,32 All these structures revealed a common binding mode for each tethered agonist, which adopted a 1.5-turn α-helical conformation with three conserved hydrophobic residues buried deep in the pocket (Fig. 2C). Conversely, before release from the extracellular GAIN domain, the Stachel peptide forms a β-sheet conformation with other strands within the GAIN B subdomain (Fig. 2B), suggesting that the Stachel peptide undergoes conformational change from an unreleased inactive state to a released active state.33,34
Nevertheless, how the Stachel peptide is released from GAIN subdomain B remains elusive. Our simulations revealed that the Stachel peptide exhibits considerable stability and shares similar dynamics compared to the surrounded strands over a timescale of 1 us35. This indicates that the Stachel peptide may not spontaneously disengage from the core of GAIN B subdomain and that a tensile force is likely needed to disrupt the interaction network. Whether typical physiological forces are sufficient to induce the release of Stachel peptides is a pivotal question that requires exploration using biophysical techniques beyond structural studies.
3. Single-molecule studies on aGPCRs
Using a single-molecule tweezer system, we and others have investigated the force response of GAIN domain-containing fragments of representative aGPCRs (Fig. 3A).36,37 Our findings demonstrate that when applying a force loading rate of 1.0 pN/s, the GAIN domains of ADGRG1 and ADGRL1 undergo structural unfolding within a force range of 5-10 pN, as evidenced by stepwise extensions of 10-30 nm. This step size indicated partial unfolding of the GAIN domain relative to the total extendable length of the loaded fragments (Fig. 3B). Subsequent to these unfolding events, the Stachel peptides dissociated from the GAIN domains within a force range of 10-20 pN in both receptors (Fig. 3C). These data indicate that unfolding precedes Stachel dissociation and that the unfolding likely occurs within the GAIN B subdomain, thereby facilitating Stachel peptide dissociation by disrupting its interactions with surrounding residues in the GAIN B subdomain. These results are consistent with the notion that without partial unfolding, Stachel peptide dissociation is less likely because of the anticipated catch-bond kinetics arising from the overall β-sheet structure of GAIN B. Thus, partial unfolding of the GAIN B subdomain may precisely disrupt the interaction network surrounding the Stachel peptide, thereby facilitating its dissociation. Moreover, our single-molecule data suggested that dissociation between the GAIN A and B subdomains likely precedes the partial unfolding of GAIN B36. Similarly, Zhong et al. found that the GAIN domain of ADGRL3/LPHN3 can be dissociated by physiologically relevant forces of 1-10 pN. Both studies support the mechanical activation hypothesis of aGPCRs, highlighting the sensitivity of the GAIN domain structure and its detachment to physiological forces. These results are consistent with a recent in vivo study suggesting that NTF/CTF dissociation occurs in vivo and that mechanical force is necessary for the activation of the neural latrophilin-type aGPCR Cirl from Drosophila.38
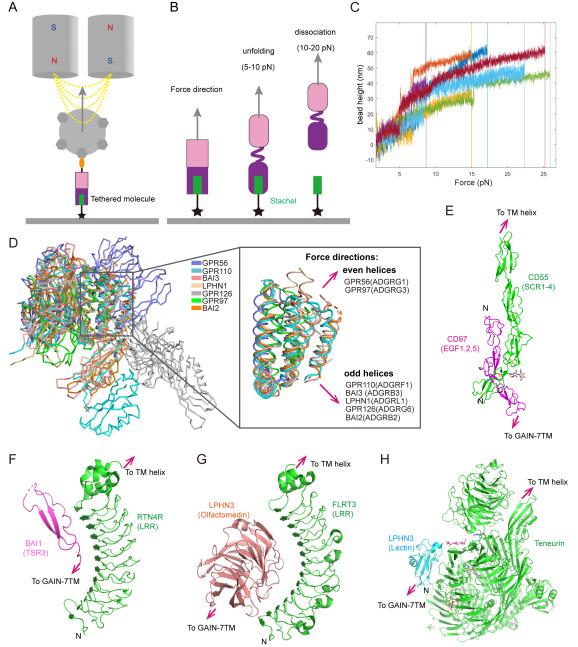
Fig. 3. Force magnitude and direction on aGPCR. |
When force is transmitted from the NTF to the GAIN subdomains, the connection between the GAIN A and B subdomains can potentially influence mechanical dissociation of the Stachel sequence. Based on the determined ECR structures, two major connecting geometries between GAIN A and B subdomains were identified (Fig. 3D): one group featured an odd number of helices in the GAIN A subdomain, with the preceding adhesion domains extending toward the cell membrane, while the other group exhibited an even number of helices in the GAIN A subdomain, with the preceding adhesion domains moving away from the cell membrane.35,39,40,41,42,43 Analysis of the sequences of GAIN domain-containing aGPCRs revealed significant diversity in the GAIN A subdomains compared to the GAIN B subdomains, with the total distribution of aGPCRs evenly split between the two groups.
These two types of connecting geometries give rise to two distinct stretching geometries at the Stachel-GAIN B interface: a near-shear-force geometry (for even helices in GAIN A) and a near-unzip-force geometry (for odd helices in GAIN A). Intriguingly, our single-molecule studies on ADGRG1 (even helices) and ADGRL1 (odd helices) revealed similar partial unfolding signals of the GAIN B domains and Stachel dissociation, despite their significantly different force geometries.35 We speculated that the application of force to the GAIN domain may initially disrupt the interaction between the GAIN A and GAIN B subdomains; therefore, the GAIN B subdomains would experience a consistent force geometry, thus leading to similar partial unfolding signals before Stachel sequence dissociation.36 Alternatively, in response to force, the N-terminal helix of the odd-helical GAIN A subdomains may be fragile and readily unfold, generating a shear force geometry similar to that of the even-helical GAIN A subdomains. In summary, our single-molecule study of the GAIN domains of ADGRG1 and ADGRL1 may have already generated a framework for understanding the force response of all aGPCRs.
The mechanism by which the force is generated in the NTF remains incompletely understood. However, the force generated by the flow-induced stretching of cells is a well-understood and extensively studied phenomenon.18 Our recent study proposed an additional plausible mechanism, suggesting that forces may arise from cell migration, potentially leading to Stachel dissociation when the link between aGPCR and the extracellular matrix is sufficiently stable.36 Other potential mechanisms of force generation include membrane dynamics resulting from actomyosin contraction at adherens junctions, which merit further investigation in future studies.
Another crucial question is how the ligand-receptor complex can withstand mechanical forces in a circulating environment. A prominent example is the CD97-CD55 complex, which has demonstrated that ADGRE5/CD97 binds to its ligand CD55 via an antiparallel binding mode, leading to a shearing-stretch geometry known to resist the force applied to the protein interface (Fig. 3E).44,45 Hence, the mechanical force generated by circulation can be transmitted to the downstream GAIN domain for unfolding and activation, as previously mentioned. Similarly, other characterized complexes between aGPCRs and their ligands (ADGRB1/BAI1TSR3-RTN4R, ADGRL3/LPHN3lectin-Teneurin, and ADGRL3/LPHN3Olfactomedin-FLRT3LRR) also exhibit opposite directions for the termini that extend to the transmembrane regions of each complex (Fig. 3F-H).43,46,47 Despite variations in the binding modes, their interfaces were roughly parallel to the potential force directions, suggesting a similar mechanism to resist the mechanical force, as indicated in the CD97-CD55 complex. Furthermore, certain β-strands on at least one side of these interfaces may contribute to stiffness in the presence of mechanical forces.
The aforementioned studies have effectively bridged the gap in understanding the mechanical activation model for these self-cleaved aGPCRs based on previous functional and structural studies. Nevertheless, approximately 1/3 aGPCRs are non-self-cleavable, prompting questions about the necessity of Stachel peptide dissociation for their mechanical activation. It has been proposed that a partially unfolded GAIN B subdomain alone is sufficient to expose and activate the Stachel sequence and achieve activation.36 Additionally, recent cryo-EM studies on a cleavage-deficient mutant of ADGRF1/GPR110 revealed that an active conformation with Stachel peptide already released from the GAIN domain and bound to the orthosteric pocket.48 Thus, regardless of the GPS cleavage status, the receptor may pre-exist in multiple states on the cell surface, including a fraction in which the Stachel peptide is already dissociated from the GAIN domain, leading to interactions with 7TM to trigger G protein coupling. This hypothesis aligns with the fact that many adhesion GPCRs display a certain level of basal activity and is supported by functional studies showing that GPS cleavage is not essential for the function of several aGPCRs.35,49,50
4. Conclusions and future prospects
In summary, the recent crystal and cryo-EM structures have provided insights into the overall structural arrangement of aGPCRs and the transition of Stachel peptide from the GAIN domain-embedded strand conformation to the 7TM-loaded α-helical conformation, supporting tether agonist engagement as the major model of aGPCR activation. Single-molecule studies have further demonstrated that physiological tensile forces are sufficient to induce unfolding of the GAIN domain and dissociation of NTF/CTF, which offers valuable insights into the mechanosensing mechanism of aGPCRs. Dysfunction of aGPCRs has been linked to various diseases and even cancers; thus, these studies provide an alternative way to regulate the function of aGPCRs using mechanical force, known as mechanotherapy. As aGPCRs can trigger divergent intracellular signaling upon sensing mechanical stimuli, they may also potentially be developed as mechanosensors.
Several outstanding questions remain to be answered regarding the detailed mechanism of aGPCRs such as: 1) Given that current structural studies suggest loose orientations between the NTF and CTF, how do the dynamics of the NTF with regard to the CTF on the self-cleaved aGPCRs compare to the non-cleavable ones? 2) What are the detailed unfolding procedures of the GAIN subdomains when a mechanical force is applied and how do the varied preceding adhesion domains affect the mechanical response of the GAIN subdomains? 3) In addition to force generation via flow perturbation and cell migration, what other mechanisms underlie force generation in the NTF? Therefore, further structural and mechanical studies at the full-length receptor scale or at the cell-based level are necessary for a comprehensive understanding of the role of mechanical activation in aGPCR signaling.
Ethics approval and consent to participate
Not applicable.
Ethical approval
This study does not contain any studies with human or animal subjects performed by any of the authors.
CRediT authorship contribution statement
Yueming Xu: Data curation, Formal analysis, Writing - original draft. Huanhuan Xu: Investigation, Resources. Jie Yan: Supervision, Validation, Writing - review & editing. Gaojie Song: Conceptualization, Supervision, Writing - review & editing.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgments
This work was supported by the National Natural Science Foundation of China (32171215) and Basic Research Program of the Science and Technology Commission of Shanghai Municipality (21JC1402400).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}