
1. Introduction
Bone tissues can be regarded as an osteoimmune system that performs locomotive functions, while the bone marrow (BM) serves as the primary site for hematopoiesis, housing hematopoietic stem cells (HSCs) and mature immune cells, such as neutrophils, macrophages, B cells, and T cells.1,2 The interaction between immune and bone cells within the BM contributes to the functioning of the “osteoimmune system.” Extensive research in osteoimmunology has provided insights into the essential interplay between bone and immune cells, which plays a critical role in the precise regulation of bone homeostasis.3 Osteoimmunology was initially established to investigate the molecular mechanisms contributing to bone degradation within the framework of inflammatory diseases. However, osteoimmunology has improved with advancements in biomaterials used in tissue engineering.
Several methods have been developed in bone regeneration to create biomaterials that mimic the natural extracellular matrix (ECM) and provide an optimal environment for bone healing. The ECM is a network-structured region that surrounds cells and provides mechanical support for forming new cells, tissue growth, and absorption of physical stress from physiological functions.4 Mechanical cues, such as surface patterns, dimensionality, stiffness, and contractility, can be detected when cells are exposed or drawn to the ECM.5 As cells push and pull on the ECM during various biological processes, the ECM initially resists these actions, governed by ECM stiffness.6 Stiffness is typically described by the elastic modulus, which ranges from hundreds of pascals to several kilopascals in various tissues. Through the control of mechanical cues such as stiffness, an increasing number of studies have shown a relationship between ECM stiffness and cell fate and activity, including proliferation and differentiation.7 According to a previous report, bone marrow mesenchymal stem cells (BMSCs) can perceive, process, and trigger mechanical cues from the ECM, thereby inducing the transcriptional regulation that governs various gene expression patterns and affects cell fate.8 Therefore, the use of mechanical stimuli directed toward BMSCs is considered for creating scaffolds in engineered tissue constructs. Stem cell-substrate interface signals can drive osteogenic differentiation, which is aided by stiff substrates and promoted by soft substrates for BMSC adipogenic differentiation.9,10 In addition, the chondrogenic differentiation of BMSCs has a significant impact on softer scaffolds by controlling the scaffold stiffness, whereas osteogenic differentiation improves on stiffer scaffolds.8,9
Directing the regulation of early immunological processes is becoming increasingly important in the domains of tissue engineering and biomaterial design as knowledge about the function of the immune system in beginning and promoting regeneration. Recently, researchers have shown that matrix rigidity plays a vital role in regulating the HSC niche. Soft BM organoids that can rejuvenate HSCs have also been developed effectively.11,12 The hematopoietic apex, where HSCs are located, is responsible for creating immune cells.13 Does this imply that the immune cells prefer a pliable matrix? Tissue or matrix stiffness is linked to the etiology and course of the disease and is a common pathological marker of immunological organs.14 Physical stiffness is an important mechanical stimulus that affects the behavior of immune cells.15 Most immune cells adapt their phenotypic traits, migration, proliferation, and immune activation in response to changes in the stiffness of their environment.16 Recently, there has been increasing focus on studying the relationship between stiffness and immune responses to determine the basic mechanisms and potential therapeutic approaches that can trigger a favorable innate immune response during in vivo implantation, promoting bone healing. Differences in the osteogenic microenvironments may suggest differences in stiffness. The extracellular matrix is not linearly elastic, and even stiffer skeletal tissues, such as bones, are viscoelastic, with loss moduli of approximately 10% of the storage moduli.17 In addition, cellular activity can lead to material degradation, which would probably change the local matrix to a more viscoelastic state.18 However, the variable and noncomparable results of immune cell responses to matrix stiffness may be confusing.
This review aims to offer a thorough and efficient understanding of cell-matrix interactions in the context of bone tissue engineering. This goal can be achieved by summarizing existing studies that have investigated the influence of substrate stiffness on immune cell responses. The initial focus of this study was to introduce different levels of ECM stiffness that are frequently utilized in tissue regeneration. Our subsequent focus was to elucidate the advancements in related research pertaining to the impact of substrate stiffness on immune cells in the context of bone regeneration. It is crucial to acknowledge that biomaterial-based scaffolds play a vital role in investigating cellular responses and their underlying molecular mechanisms and uncovering the importance of substrate stiffness in both physiological and pathological processes. This review aims to provide valuable insights and contribute to future advancements in biomaterial-based scaffolds in regenerative medicine.
2. ECMs with different stiffnesses attributed to native tissues
The ECM functions as a cellular microenvironment, enabling the transfer of external mechanical forces and internal mechanical stimuli to resident cells. Moreover, it influences the fate of these cells by regulating their proliferation and specialization. Numerous studies have provided evidence for the significant influence of matrix stiffness, including both natural ECM and engineered substrates, on cellular behavior and tissue formation in tissue engineering.3,6,19
Resident cells control the flexibility and molecular makeup of the matrix, thereby regulating the ECM.20 The stiffness of the ECM varies significantly owing to the fundamental characteristics of its constituent molecules and their distinct self-assembled hierarchical designs. Finite element investigations have demonstrated that Young's modulus of brain parenchyma is approximately 584 Pa.21,22 Additionally, Segel et al. used atomic force microscopy (AFM) to confirm that the prefrontal cortex gradually stiffens with age and showed that the primary regulator of “cellular activity” of multipotent stem cells in the central nervous system is ECM stiffness rather than cell age or ECM chemistry.23 Using AFM, the compression elasticity of heart muscle fibers in cows varied between 23 and 42 kPa in both the rigor and relaxing states.24 The Young's modulus of native human lungs has been reported to be approximately 1.9 kPa, but decellularized human lungs were found to have a similar value of 1.6 kPa.25 The elastic modulus of human liver samples measured using AFM varied between 0.18 and 0.02 MPa.26 Using AFM, Young's modulus of the decellularized rat pancreas was approximately 3.7 kPa.27
The bone is a tissue with a rich blood supply that undergoes ongoing remodeling. In vitro research, promoting osteogenic differentiation often involves using materials such as collagen, hyaluronan, mineralized calcium, fibrin, or their combinations.28 By manipulating the elasticity of gel matrices to fall within the range of 25-40 kPa, which corresponds to the collagenous osteoid matrix (a precursor to rigid calcified bone with an elastic modulus of 1 GPa), mesenchymal stem cells can be prompted to differentiate into osteoblasts.29,30 The elastic modulus of human cancellous bone typically falls within the range of 1.3-7.8 GPa, while that of cortical bone is greater, ranging between 12 and 20 GPa.31,32 These values are significantly higher than those observed in the osteoid matrix and differ by several orders of magnitude.
The stiffness of tissues varies significantly. To ensure that the stiffness of biomimetic materials is comparable across studies, the measurement methods should first be standardized. Currently, the commonly used methods for measuring the stiffness of biological and material samples can be distinguished at the macro and micro levels. Macroscopic characterization, that is, the bulk modulus, can be acquired from stress-strain measurements using a universal testing machine. The bulk modulus is related not only to the longitudinal stress-strain relationship of the material but also to its compressibility and Poisson's ratio.33 Although widely applied in research, this approach can be problematic for materials such as hydrogels that contain large amounts of water and are nearly incompressible.34,35 This makes the bulk modulus highly sensitive to water content, which does not accurately represent material stiffness. From the perspective of microscopic characterization, liquid-phase AFM and nanoindentation can reflect what the cells sense.36,37 AFM is a versatile methodology capable of investigating the physical properties of complex biological tissues under physiologically relevant conditions without requiring sample chemical manipulation, with low to high forces in a localized region or from slow to fast temporal stimulation.38 Owing to its high versatility, AFM has become a leading biophysical tool for determining the physical properties of complex biological specimens, including cells and tissues, in relevant physiological environments. Thus, AFM might be a better choice for measuring stiffness.
3. The effects of matrix stiffness on various immune cells in bone biology.
In this section, we explore the effects of matrix stiffness on various immune cells in the bone. These matrices include natural and synthetic ECMs (hydrogels).6 Thus, despite the different names used in various studies, the stiffness difference was distinguished by numerical magnitude. For clarity, the results are summarized in Table 1.
Table 1. The effects of matrix stiffness on various immune cells in bone biology. |
| Immune cells | Authors | Culture system | Magnitude of stiffness (measure methods) | Regulatory pathway | Sub-type/state of activation |
|---|---|---|---|---|---|
| Neutrophil | Abaricia et al.43 | 2D | 0.2-32 kPa (Not clear) | Integrin/FAK | Stiffer - NETs |
| Jiang et al.44 | 3D | 9-55 kPa (AFM) 75-110 kPa (Nanoindentation) 1.5-5.7 kPa (Compressive test) | JAK1/STAT3 | Stiffer - N2 type | |
| Macrophage | Chen et al.34 | 2D | 2.6-63.5 kPa (Compressive test) | NF-κB signaling | Stiffer - M2 type |
| Zhuang et al.35 | 2D | 2-29 kPa (Compressive test) | Not clear | Stiffer - M1 type | |
| Okamoto et al.57 | 2D | 4-100 kPa (AFM) | PPARγ signaling | Softer - M2 | |
| He et al.59 | 3D | 1.6-60.5 kPa (Compressive test) | Not clear | Stiffer - M1 type | |
| B cell | Shaheen et al.61 | 2D | 2.6-22.1 kPa (Not clear) | PKCβ/FAK | Stiffer - activation |
| Wan et al.63 | 2D | 2.6-22.1 kPa (AFM) | Not clear | Stiffer - activation | |
| T cell | Judokusumo et al.74 | 3D | 10-200 kPa (Compressive test) | downstream of T-cell receptor | Not clear |
| Saitakis et al.76 | 2D | 0.5-100 kPa (Tensile elastic modulus) | Not clear | Stiffer - increased metabolism | |
| Chin et al.79 | 2D | 7.1-50.6 kPa (AFM) | Not clear | Stiffer - more IL2 | |
| O'Connor et al.80 | 2D | <100 kPa (custom-built indentation apparatus) | Not clear | Softer - more IL2 | |
| Shi et al.82 | 2D | 7.5-140 kPa (indentation) | AMPK/oxidative phosphorylation | Stiffer - more Treg cells | |
| Chirivì et al.83 | 3D | 0.4-2.5 kPa (Compressive test) | Not clear | Stiffer favoring CD4+ T cells over CD8+ T cells |
AFM, atomic force microscope. FAK, focal adhesion kinase. JAK, janus kinase. STAT, signal transducer and activator of transcription. NF-κB, nuclear factor kappa-light-chain-enhancer of activated B cells. PPAR, peroxisome proliferator-activated receptor. PKC, protein kinase C. AMPK, AMP-activated protein kinase. NET, neutrophil extracellular trap. IL, interleukin. Treg, regulatory T cells. |
3.1. Neutrophils
Neutrophils (Neus), which serve as the initial defense mechanisms within the innate immune system, play vital roles in wound healing and immunity. These cells predominantly govern the early inflammatory response and serve as primary stimulators of the regenerative process.39 Specifically, anti-inflammatory N2-Neus express various anti-inflammatory signals that attract mesenchymal stem cells and convert proinflammatory immune cells into anti-inflammatory cells. In contrast, the early inflammatory response is triggered by traditionally proinflammatory N1-Neus, which stimulates phagocytic cells to eliminate damaged tissues.39,40,41 N2-polarized composite hydrogel scaffolds were recently developed by adding growth factors generated by N2-Neus to hydrogels to reduce inflammation and enhance angiogenesis.42 To optimize the number of angiogenic therapeutic options for ischemic disorders and bone abnormalities, it would be helpful to investigate how hydrogels with different matrix mechanical conditions alter Neus phenotypes.
Neus activation may be regulated by substrate stiffness43 (Fig. 1). The expression of cytokines and chemokines did not change significantly when Neus were cultured in two-dimensional (2D) systems at less than 8 kPa (measured by a compressive test for bulk stiffness).43 However, our previous study showed that within a limited range of stiffnesses (1.5-5.7 kPa), distinct Neus phenotypes might be evoked in the three-dimensional (3D) culture system.44 A denser matrix enhanced the development of the N2 phenotype. Conventional indicators, namely CD182 for N2-Neus and CD54 for N1-Neus, were evaluated based on the behavior of Neus within the tumor microenvironment.45 Neus cultured in a more rigid matrix exhibited increased tumor growth factor beta (TGFβ) receptor and CD182 expression. Neus grown in hydrogels with low stiffness produced more reactive oxygen species (ROS) and released more interferon gamma-induced protein 10 and tumor necrosis factor-alpha (TNFα), whereas those grown in hydrogels with high substrate stiffness presented high gene expression levels of CD206 and C-X-C motif chemokine ligand 12 and secreted more interleukin (IL)-10, indicating a propensity toward N2 polarization.45 The local stiffness of the hydrogels, as assessed using AFM, varied in the range of 9-55 kPa.44 Neutrophil polarization was initially observed in the local microenvironment of cancer, and the findings indicate that substrates with stiffnesses varying from 1 to 50 kPa (evaluated using AFM) may accurately reproduce this environment.46,47
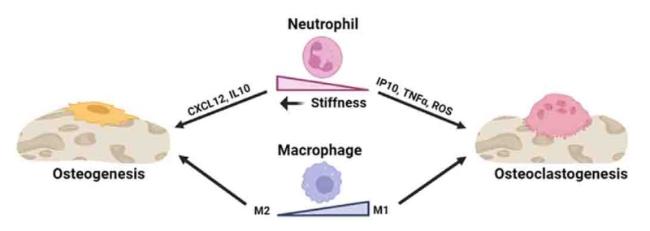
Fig. 1. The effects of stiffness on neutrophils and macrophages in the aspect of bone remodeling. |
Furthermore, net-like structures of decondensed chromatin, antimicrobial proteins, and enzymatic activators of cytokine precursors are released by neurons during neutrophil extracellular trap (NET) synthesis.48 Nuclear and cell membrane rupture results in the formation of NETs. Reportedly, there is a stiffness-dependent increase in NET production; in a 2D system, stiffness levels greater than 8 kPa may significantly increase NET formation.43,49 It is simpler to induce NET growth with the same stiffness as in the 3D culture technique than with the 2D culture system.50 Additionally, a long-term study indicated that the development of NETs is a crucial factor in the aftermath of the “inflammaging” phenomenon.51
3.2. Macrophages
Macrophages, which originate from a monocytic lineage and include osteoclasts, are integral components of the innate immune system.52 These cells are widely distributed throughout different body tissues and primarily function in phagocytosis, engulfment, and digestion of cellular debris and foreign substances in response to infection or tissue damage.53 Following the introduction of a biomaterial, immune cells promptly migrate to the implantation site and initiate localized inflammatory reactions.54 Although multiple cell types are involved in the inflammatory response, macrophages play a critical role by secreting chemokines and cytokines that directly affect other cells.
Pioneering studies have elucidated the multifaceted nature of macrophages, encompassing both classically activated M1 and alternatively activated M2 subtypes.55 M1 macrophages are commonly associated with generating proinflammatory cytokines, phagocytosis of microbes, and initiating immune responses.56 Conversely, M2 macrophages are primarily involved in wound healing and tissue repair through the secretion of anti-inflammatory cytokines, remodeling activities, and proangiogenic factors.56 Hence, the modulation of macrophage behavior is useful for regulating inflammatory responses and promoting the integration of biomaterials and medical devices into tissues.
Macrophages can modify their morphology in response to alterations in substrate stiffness (Fig. 1). This phenomenon was also observed when macrophages were cultured on soft hydrogels, resulting in a more elongated form than macrophages cultured on rigid hydrogels, as indicated by a reduced minor axis.35 Moreover, macrophages can promptly adapt to their phenotype and polarization state in response to localized stimuli, including physical signals. One study revealed that hydrogels with a stiffness of 29.2 kPa effectively induced M1 macrophage polarization, as evidenced by elevated secretion of TNF-α and IL-6 and reduced secretion of TGF-β and C-C motif ligand 17 following a 3-day culture period.35 Additionally, previous studies have consistently demonstrated that decreased substrate stiffness promotes M2-like macrophage activation, consistent with the findings of other studies.57,58 Moreover, previous studies examined the influence of matrix stiffness on the osteogenic capacity of BMSCs, specifically exploring the role of macrophages in co-cultures to modify the effects of matrix stiffness on BMSCs.59 In contrast, macrophages residing within high-stiffness gels exhibited an increased inclination toward polarization toward the proinflammatory M1 phenotype. By employing either conditioned medium-based incubation or transwell-based co-culture, macrophages encapsulated within a low-stiffness matrix were shown to benefit the osteogenesis of co-cultured BMSCs. Conversely, macrophages residing within high-stiffness gels adversely influence the osteogenic differentiation of cells.59 However, other studies have reported contradictory results. After confirming the biocompatibility of the hydrogels, experiments were conducted using bone marrow-derived macrophages (BMMs) from mice incubated with different hydrogels. When exposed to simulated low substrate stiffness (2.55 ± 0.32 kPa), the BMMs exhibited increased expression of CD86 on their cell surface, increased production of ROS within the cells, and increased secretion of IL-1β and TNF-α in the supernatant. In contrast, BMMs exhibited heightened CD206 expression, decreased ROS production, and enhanced secretion of IL-4 and TGF-β when exposed to medium stiffness (34.88 ± 4.22 kPa).34 The disparate outcomes observed can be attributed to using different substrate stiffness levels in the investigations.
This suggests that matrix stiffness affects macrophage behavior and macrophage-mediated inflammatory responses. These findings provide significant insights for advancing immunology-informed biomaterials to promote tissue regeneration by modulating the inflammatory response mediated by macrophages.
3.3. B cells
B cells play a major role in the immune response by secreting antibodies specific for antigens (Ags) and offering a sustained defense against invading infections. They also serve as antigen-presenting cells, release cytokines that are both pro- and anti-inflammatory and influence other immune response-related cells, such as T cells, both positively and negatively.60 B cells recognize ECM-associated Ags in a contact-dependent manner and via the B cell receptor when the ECM stiffness of the tissue varies significantly from 0.012 to 20 kPa.61 According to recent studies, B cell activation is effectively regulated by the degree of stiffness of the substrate while delivering Ags.62 When Ags are attached to highly rigid substrates, B cells react significantly more strongly, as evidenced by the build-up of B cell receptors, phospho-spleen tyrosine kinase, and phosphotyrosine molecules within the B cell immunological synapse.63 B cells cannot distinguish between substrates with different stiffnesses unless protein kinase C beta-dependent focal adhesion kinase (FAK) is activated, after which FAK subsequently increases B cell adhesion and spreading responses.61,64
Additionally, B and bone cells exhibit complex and intimate interactions. B cells release cytokines, which are linked to maintaining bone homeostasis during inflammation.65 It has been reported that B cells produce proinflammatory cytokines and receptor activators of nuclear factor-κ b ligand (RANKL), which can drive osteoclast development and alveolar bone resorption.60 In addition, B cells produce RANKL in patients with periodontitis. According to one study, 90% of B cells are RANKL-positive, which is greater than the corresponding number of T cells.66 Mature B cells do not produce osteoclasts; they indirectly affect osteoclastogenesis by producing RANKL or the osteoclastogenesis inhibitory factor. The effects of these cytokines on osteoclastogenesis are noteworthy because they regulate osteoclast formation. At that point, as shown in conditions marked by bone degradation, such as inflammatory arthritis and periodontal disease, B cells and osteoclasts contribute to the pathophysiology of the disease.67 Conversely, memory B cells inside the periodontal tissues of patients with periodontitis may exhibit elevated TGF-β1 expression and impede BMSC development into osteoblasts.68 The osteoblastogenesis of BMSCs is reduced by activating B cells through Notch signaling, and the osteoblastogenesis of BMSCs is partially rescued by the suppression of B cells or inactivation of Notch signaling.69 Different stiffness ranges may be responsible for these varying findings.
Research has been conducted on the effects of substrate stiffness on B cells and the relationship between B cells and bone remodeling61 (Fig. 2). However, the impact of stiffness on B cell function in osteoimmunology has not received much attention. This could mean that the riddle of creating appropriate materials for tissue engineering by controlling osteoimmunology remains unresolved.
Fig. 2. Stiffer substrates promote lymphocytes cells activation. |
3.4. T cells
T cells, which are derived from hematopoietic cells and undergo maturation in the thymus, exhibit distinct surface receptors and functions that are contingent on their differentiation stage.70 CD8+ cytotoxic T cells can directly eliminate infected or mutated cells. Conversely, CD4+ T helper (Th) cells interact with other immune cells, such as B cells, through surface receptors and cytokine secretion, thereby modulating their activation state. Th cells can be further classified based on cytokine expression patterns.71 Th1 cells secrete interferon-gamma, IL-2, and TNFα, primarily influencing macrophages. TNFα facilitates the upregulation of RANKL expression by macrophages, thereby stimulating osteoclastogenesis. Th2 cells secrete various interleukins, including IL-4, IL-5, IL-9, IL-10, and IL-13, primarily influencing B cells, mast cells, and granulocytes. Another distinct subset of T cells, known as Th17 cells, can be identified by their secretion of IL-17.72 Additionally, CD4+CD25+ Tregs, called regulatory T cells, play a crucial role in modulating the immune system and preserving immune tolerance.73
With the advancement of immunotherapies, the mechanosensing abilities of T cell receptors have attracted increased attention.74 New research has emerged concerning how mechanosensitivity allows for the adjustment of crucial immune functions, such as T cell activation and death of target cells by cytotoxic T lymphocytes.75 Saitakis et al.76 demonstrated that T cells can distinguish a large range of stiffness values in the body and modify their responses appropriately. Specifically, within the physiological range of 0.5-100 kPa, T cell migration and morphological alterations are influenced by stiffness. Moreover, T cell metabolic characteristics and cell cycle progression increased only at the maximum stiffness examined (100 kPa). The cellular response of T cells can be either biphasic or monotonous, depending on the triggered surface receptors.77,78 T cells react differently to stiffness in the environment when they adhere only through the T cell receptor complex, reaching maximum spreading on an ideal substrate stiffness similar to that of skilled antigen-presenting cells. However, this biphasic response is disrupted in the presence of extra integrin leukocyte function-associated antigen 1 ligands, and cell spreading increases steadily with stiffness up to a saturation point.77 Taking advantage of the fact that T cell activation can be largely affected by the stiffness of the matrix, Chin et al.79 reported the possibility of developing immunostimulatory biomaterials that are mechanically adjusted to balance the stimulatory intensity and therapeutic T cell capacity for downstream proliferation. In this study, IL-2 was quantified by evaluating the stiffness of hydrogels from 7.1 ± 0.4 to 50.6 ± 15.1 kPa. Compared with stiffer substrates, softer substrates elicit a four-fold increase in the production of IL-2 and proliferation of human CD4+ and CD8+ T cells.80 Tabdanov et al.81 engineered nanotextured elastic platforms for managing T cell migration, based on which useful tactics could be created to boost the effectiveness of immune therapies. Shi et al.82 suggested that Treg induction on polyacrylamide gels increases with increasing material stiffness and is influenced by the elastic modulus of the substrate. Mechanistically, oxidative phosphorylation is more common during Treg induction on stiff substrates. While the activation of AMP-activated protein kinase boosted Treg induction in the softer sample but not in the harder sample, the inhibition of ATP synthase greatly decreased the rate of Treg induction and eliminated the differences among gels.
Advancements in emerging technologies have contributed to a better understanding of the involvement of T cells in bone tissue engineering. Simultaneously, extensive research has been conducted to investigate the influence of substrate stiffness on T cell functionality.82,83 Nevertheless, the precise implications within the realm of osteoimmunology are yet to be fully elucidated (Fig. 2).
3.5. The mechanism by which immune cells detect alterations in matrix stiffness
In general, cells interact with their environment through integrins, which are transmembrane proteins composed of alpha and beta subunits. Integrins function as membrane receptors, recognize ECM protein domains, and facilitate signal transduction from the ECM to the cells.84 It has been reported that blocking integrins alters neutrophil migration and function, as they play a fundamental role in neutrophil adhesion.85 Furthermore, neutrophils sense and respond to physical cues through integrin/FAK signaling, and phosphoinositide 3-kinase inhibition reduces neutrophil cell area on polyacrylamide substrates.43,86 For macrophages, stiffness-dependent changes in cell function reportedly require Piezo1, and M2 macrophages are activated by modulating the ROS-initiated NF-κB pathway.34,87
In lymphocytes, the T cell receptor acts as a mechanosensor.88 T cells adapt to the stiffening of artificial antigen-presenting cells by changing the loading rate of their pulling forces.62 B cell mechanosensing-governed activation also requires B cell receptor signaling molecules.61 Additionally, B cells discriminate substrate stiffness in a protein kinase C beta-mediated FAK activation-dependent manner.61
4. Conclusions and future prospects
Previous research has revealed a significant correlation between substrate stiffness and immune cell response, which is highly important in bone tissue regeneration guided by substrates. A thorough understanding of the complex mechanical interactions between cells and their ECM will assist in advancing tissue engineering strategies to direct cell fate, enhance tissue regeneration, and enable the construction of tissue engineering platforms capable of monitoring diverse biological processes involved in bone regeneration.
First, the range of approaches available to exclusively regulate the substrate stiffness is somewhat constrained. Modifying the matrix stiffness is typically accompanied by alterations in additional biophysical and biochemical stimuli, such as architectural or surface chemical factors. Consequently, the fabrication of biomimetic materials that solely incorporate stiffness variations can significantly enhance our understanding of the regenerative processes induced by materials. Moreover, there are notable disparities between the outcomes obtained from 3D models and those derived from 2D models. The accurate reproduction of the complex three-dimensional mechanical milieu of immune cells within substrates remains a distinct obstacle that necessitates further investigation and consideration in the future. Furthermore, while research on cellular responses in engineered tissues with varying substrate stiffnesses has offered insights into how immune cells react to different stiffness levels for bone regeneration, a comprehensive understanding of the underlying mechanism governing stiffness-mediated bioactivities within immune cells at different stages is yet to be fully elucidated. Additional research is necessary to comprehensively elucidate the complexity of this mechanical feedback loop between cells and the ECM, particularly considering the influence of intercellular interactions.
Author contributions
TJ and NJO designed the review. TJ, MTZ, and RML drafted the manuscript, and NJO revised the manuscript. All the authors have approved the manuscript.
Ethical approval
This study does not contain any studies with human or animal subjects performed by any of the authors.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgments
This work was supported by the National Natural Science Foundation of China, China (82101047) and the Fundamental Research Program funding of the Ninth People's Hospital affiliated with Shanghai Jiao Tong University School of Medicine, China (JYZZ138).

{kind=link}
{kind=link}
{kind=link}
{kind=link}