
1. Introduction
Colon cancer is one of the most common malignant tumors and ranks second in terms of cancer-related deaths worldwide.1,2 However, distant metastasis is considered the primary cause of clinical treatment failure in patients with colon cancer,3,4,5 with an overall 5-year survival rate accounting for only 13.3%.6 Our bioinformatics analyses found that the higher expression levels of yes associated protein 1 (YAP1), which is a core transcriptional coactivator and transducer of mechanical stimuli in patients with colon cancer, were associated with their metastasis and poor prognoses. Physical and chemical factors in the solid tumor microenvironment are key to regulating the occurrence and development of cancer cells.7,8 An increasing number of researchers have focused on the mechanochemical coupling mechanism of the effects of increased extracellular matrix (ECM) stiffness, blood flow/interstitial flow shear stress, and static pressure on tumor cell proliferation and metastasis.9,10 ECM stiffness plays an important role in the regulation of cancer cell migration and invasion.11,12 Cells can sense and respond to various extracellular mechanical stimuli to ensure proper physiological functions, such as adhesion, migration, invasion, and progression of various diseases, including cancer.13,14 Extracellular mechanical signals are transmitted by activating intracellular signaling pathways via mechanical sensors in the cell plasma membrane.15,16 The force applied to ECM-integrin adhesion can activate focal adhesion kinase (FAK) and promote the formation of focal adhesions (FAs), thereby recruiting adhesion-related proteins (e.g., vinculin, talin, and paxillin) to trigger downstream signaling cascades.17 FAs are dynamic multi-protein complexes that participate in ECM adhesion and play an important role in translating ECM stiffness signals into intracellular chemical responses. The dynamic cycle of FAs assembly and disassembly at the edge of a migrating cell provides a directional force for movement; FAs dysfunction is an important step in tumor cell invasion and metastasis.18 Therefore, it can be hypothesized that colon cancer cells (CCCs) sense and respond to changes in ECM stiffness via YAP1 and FAs, leading to invasion and metastasis.
To verify this hypothesis, we used a polyacrylamide (PA) hydrogel to simulate the stiffness microenvironment of normal colon tissue and colon cancer tissue under physiological and pathophysiological conditions and investigated the effects of different ECM stiffnesses on the mechanophenotypes, including morphology, adhesion, invasion, electrophoresis rate, membrane fluidity, and Young's modulus of CCCs. In addition, we explored the molecular mechanisms by which ECM stiffness regulates the migration and invasion of colon cancer cells. Therefore, it is important to understand the mechanism of CCCs metastasis from the perspective of mechanobiology to optimize and personalize clinical treatment strategies for patients.
2. Material and methods
2.1. Bioinformatic analysis
The UALCAN database (https://ualcan.path.uab.edu)19 was used to analyze the mRNA levels of YAP1 genes in pan-cancer based on The Cancer Genome Atlas (TCGA) database. In this study, we used this database to analyze the expression of YAP1 in colon cancer and normal tissues, as well as node metastasis status. The Gene Expression Omnibus (GEO) database (https://www.ncbi.nlm.nih.gov/geo/) was used to analyze the expression levels of YAP1 in colon cancer (GSE 39582) based on distant metastasis status, and patients were subgrouped into no distant metastasis (M0) and distant metastasis (M1) groups. GEPIA (http://gepia.cancer-pku.cn)20 was used to analyze the correlation between the expression of YAP1 and paxillin in colon adenocarcinoma from TCGA database. The Human Protein Atlas (HPA) database (www.proteinatlas.org) was used to analyze the protein levels of YAP1 in colon normal and cancer tissues by immunohistochemical (IHC) staining.
2.2. Cell culture
The CCCs (HCT116) were donated by the Institute of Oncology, Affiliated Tumor Hospital of Guangzhou Medical University (Guangzhou, China). This protocol was approved by the Human and Animal Ethics Committee of Guizhou Medical University (No. 2023-46). The cells were cultured in RPMI-1640 medium (Gibco) containing 10% fetal bovine serum (Biological Industries) and 1% antibiotics (streptomycin and penicillin, Gibco). Cells were incubated in a humidified incubator at 37 °C containing 5% CO2. The cells were harvested using trypsin-EDTA (Gibco) and cultured on hydrogel substrates for further experiments.
2.3. Preparation of polyacrylamide hydrogel
PA hydrogels of different stiffnesses were prepared according to the protocols of Tse21 and Peng.22 Briefly, 40% acrylamide (Aladdin) and 2% bis-acrylamide (Sigma-Aldrich) were mixed in varying proportions, and then 10% ammonium persulfate (APS, Aladdin) and 0.1% tetramethylethylenediamine (TEMED, Sigma-Aldrich) were gently mixed and added to a hydrogel mold, which was polymerized at room temperature for 20 min to obtain a final thickness of 0.75 mm. The surface of the hydrogel substrate was applied with 0.2 mg/ml sulfosuccinimidyl-6-(4′-azido-2′-nitrophenylamino)-hexanoate (Sulfo-SANPAH, Thermo-Fisher Scientific) under 365 nm ultraviolet (UV) light for 25 min. These hydrogels were coated with collagen I (0.25 mg/ml) in phosphate-buffered saline (PBS) and incubated overnight at 4 °C. The following day, the collagen I solution was removed from the hydrogel, which were then kept in PBS at 4 °C until cell culture.
2.4. Rheological testing
A rheometer was used to determine the Young's modulus of the PA hydrogels. The detection method and calculation of the Young's modulus were based on the work published by Sana et al.23 Before measurement, the hydrogels were immersed in deionized water to remove non-crosslinked monomers, swollen to an equilibrium state at room temperature for 24 h, and then split into small round pieces with a diameter of 20 mm. Storage modulus (G′) was measured by modular compact rheometer (Anton Paar, Germany) with a 20 mm upper parallel plate geometry, oscillatory frequency sweep test from 1 to 10 Hz, and a constant 2% strain. Young's modulus was calculated using the following equation:
$\mathrm{E}=\mathrm{G}^{\prime} 2(1+v)$
where E is Young's modulus, G′ is storage modulus and v is the Poisson's ratio (0.5 for PA hydrogel).
2.5. Cell morphology
HCT116 cells were cultured on PA hydrogels at a density of 1 × 104 cells/well. After 24, 48, and 72 h, cells were photographed under an inverted microscope. Cell morphology (cell area, circularity, and aspect ratio) was analyzed using ImageJ software.
2.6. Spheroid spreading assay
The spheroid spreading assay was performed as described previously.24,25 In brief, the wells of a 96-well-plate were coated with 50 μl of 1% agarose (Millipore Sigma) in PBS, then seeded with 100 μl of the cell suspension (3 × 103 cells). Multicellular spheroids were obtained after culturing for 4 days. Then, the spheroids were collected and inoculated on cell culture plates with PA hydrogels, and images were captured at 4 and 24 h time points using an inverted microscope. Finally, to quantify the degree of dispersion, images were analyzed using ImageJ software to outline the periphery of the aggregate, and the total area of the 24 h image was divided by the area of the initial image.
2.7. Time-lapse imaging
For free migration experiments, the cells were seeded onto the PA hydrogel for 12 h. The complete medium was replaced with medium containing 1% serum. Subsequently, the live cell workstation was debugged and real-time photo parameters were set to capture images every 10 min using a 10× objective lens. The recording lasted for 9 h. The acquired images were used to generate cell trajectories using the ImageJ software. In addition, the DiPer open-source program was used to calculate the cell trajectory, average cell velocity, and mean azimuth movement.26
2.8. Atomic force microscopy (AFM)
The Young's modulus of the cells were measured using atomic force microscopy (AFM; JPK NanoRacer, Bruker). Cells were inoculated onto hydrogels of different stiffness and cultured for 48 h. Subsequently, the medium was aspirated and the cells were washed twice with PBS. The probe was then subjected to detection at a constant velocity of 2.0 μm/s, while experiencing a force ranging from 0.1 to 1 nN. Standard silicon nitride cantilever with an elastic coefficient of 0.03 N/m was employed. The probe itself possessed an elastic coefficient of 0.08 N/m, a spherical needle measuring 10 μm, and a Poisson's ratio of 0.5. The experiment was carried out under controlled conditions, specifically in a quiet and clean environment with a constant temperature of 25 °C. Forty cells were randomly selected from each sample and each cell was measured twice. Following the acquisition of force and distance curves, the JPK image processing software was utilized to fit and analyze the curves within the range of 0-200 pN in the needle loading curve. Subsequently, Young's modulus of the cells were determined based on the fitting curves.
2.9. Electrophoretic mobility (EPM)
The measurement technique employed for the EPM was outlined in a previous study.27 In brief, cells were cultured on hydrogels of different stiffness for 48 h. Subsequently, the cells were digested using trypsin-EDTA, harvested, and fixed with 1% glutaraldehyde at room temperature for 10 min. The resulting cell suspension was prepared at a concentration of 5 × 105 cells/ml using 9% (w/v) sucrose solution. The cell suspension was then injected into a cell electrophoretic migration chamber, which was mounted onto a microscope platform. During the examination using a low-power microscope, it was determined that the static layer occupied 1/10 of the distance between the front and back wall lines. Consequently, the experimental measurements were conducted within the static layer. Each group selected 20 cells and recorded the time it took for these cells to traverse two cells, with each cell measuring 50 μm. The measurements were conducted at a temperature of 25 °C and a voltage of 50 V. The average time for 20 cells was calculated and the electrophoresis rate of the cells was determined.
2.10. Cell membrane fluidity
Cells were cultured on hydrogels of different stiffness for 48 h. After washing twice with PBS, they were treated with a 1 ml solution of the DPH probe, ensuring that light sources were avoided, and the cells were subsequently incubated at a temperature of 37 °C for a period of 30 min. Following two washes with PBS, trypsin digestion was performed, followed by washing with PBS, and finally collection. The cell count was determined and the cells were suspended in 1 ml of PBS, resulting in a cell concentration of 1 × 106 cells/ml. The fluorescence intensity (I) of cells at various polarization angles was measured using a time scan on a fluorescence spectrometer. The excitation wavelength (λ EX) was set to 360 nm, the emission wavelength (λ EM) to 430 nm, and the emission slit to 5 nm. The scanning process lasted 10 s, and each sample was scanned three times. The labeled cells were measured within a maximum time frame of 2 h, ensuring cell viability throughout the measurement procedure. The calculation of fluorescence polarization (P) is determined by the formula: P=(IVV-GIVH)/(IVV + GIVH), whereby the P value exhibits an inverse relationship with the fluidity of cell membrane.
2.11. Immunofluorescence of focal adhesion
The cells were cultured on hydrogels of different stiffness for 48 h and fixed with 4% paraformaldehyde for 15 min. The cells were permeabilized with 0.5% Triton X-100 for 5 min. The cells were then washed with PBS and blocked with a 1% BSA solution for 60 min. The cells were incubated with primary antibodies (1:100), including anti-Paxilin (Abcam), anti-Talin (Abcam), and anti-Vinculin (Abcam) overnight at 4 °C. After the cells were washed thrice with PBS and incubated with a secondary goat anti-rabbit IgG (1∶200) (Abcam), nuclear counterstaining was performed using DAPI. Finally, the fluorescence was observed and photographed using a laser confocal microscope.
2.12. YAP1 knockdown of CCCs
YAP1-shRNA and shNC lentiviruses were purchased from Shanghai GENECHEM Company. Lentiviral infection was performed according to the manufacturer's protocol. Briefly, CCCs were seeded in 6-well cell culture plates at a density of 3.5 × 105 cells/well and incubated for 12 h. Then, lentivirus-containing cell culture medium was added to the cells. After 8 h, the medium was replaced with fresh medium. After 24 h, the cells were replaced with fresh complete medium and treated with 4 μg/ml puromycin. Finally, the knockdown efficiency was confirmed by qRT-PCR, western blotting, and immunofluorescence in cells, which were used for subsequent experiments. The detailed target sequences were as follows:
| Gene name | Sequence |
|---|---|
| shNC | TTCTCCGAACGTGTCACGT |
| shYAP1-1 | GCTCATTCCTCTCCAGCTTCT |
| shYAP1-2 | GCAGCAGAATATGATGAACTC |
| shYAP1-3 | GCTGCCACCAAGCTAGATAAA |
2.13. RNA isolation and quantitative real-time polymerase chain reaction (qRT-PCR)
Total cellular RNA was extracted using Trizol Reagent (Invitrogen) according to the manufacturer's protocol. cDNA was synthesized using a cDNA Reverse Transcription Kit (Thermo Fisher Scientific), and mRNA expression was analyzed by quantitative real-time reverse transcription polymerase chain reaction with SYBR green (Takara). The 2−ΔΔCt method was used to analyze the relative mRNA levels of target gene. GAPDH was used as an internal control. The following primer sequences were used:
| Gene name | Sequence |
|---|---|
| YAP1-forward | AGGAGAGACTGCGGTTGAAA |
| YAP1-reverse | CCCAGGAGAAGACACTGCAT |
| GAPDH-forward | GACCTGACCTGCCGTCTA |
| GAPDH-reverse | AGGAGTGGGTGTCGCTGT |
2.14. Western blotting (WB) analysis
Cells were cultured on hydrogels of different stiffnesses in 6-well cell culture plates and lysed on ice for 10 min using RIPA protein lysis buffer (containing PMSF, protease inhibitors, and phosphatase inhibitors). The lysates were collected in 1.5 ml centrifuge tubes and centrifuged at 12000 rpm for 25 min at 4 °C. The supernatants were collected in new prechilled 1.5 ml centrifuge tubes and stored on ice until use. The protein concentration was determined by BCA protein detection kit. Proteins were separated by 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred to PVDF membranes (Millipore). The membranes were blocked using 5% BSA at room temperature for 1 h and incubated with specific primary antibodies (1∶1000) overnight at 4 °C. After washing with TBST, the corresponding horseradish peroxidase-labeled secondary antibodies (1∶5000) were added and incubated at room temperature for 1 h, and the signal was detected using the ECL method. GAPDH was selected as the internal reference and ImageJ software was used for gray value analysis.
2.15. Statistical analysis
Data are presented as mean ± standard deviation (SD). Comparisons between two groups were performed using the unpaired t-test, and comparisons between three or more groups were performed using one-way ANOVA. P values for statistical significance are represented as ns p > 0.05, ∗p < 0.05, ∗∗p < 0.01, ∗∗∗p < 0.001, and ∗∗∗∗p < 0.0001.
3. Results
3.1. Higher YAP1 expression levels in colon cancer were associated with their metastasis and poor prognosis
Various extracellular mechanical factors regulate cancer development and progression. YAP1 is an important mechanotransducer that senses mechanical stimuli28 and translates signals to control transcriptional programs by translocating to the nucleus.29 To explore the role of YAP1 in the progression and metastasis of colon cancer, we first analyzed the expression of YAP1 in pan-cancer cells using the UALCAN online tool to obtain RNA-seq data from TCGA database. As shown in Fig. 1A and B, the mRNA expression level of YAP1 was significantly elevated in colon cancer tissues compared to normal controls. We also analyzed the YAP1 protein levels using immunohistochemical staining data from the HPA database. Compared to normal colon tissues, the protein expression level of YAP1 was significantly upregulated in colon cancer tissues (Fig. 1C), which was consistent with the corresponding mRNA levels. The relationship between the expression of YAP1 and patient clinicopathological variables along histological subtypes (Fig. 1D) and clinical stage (Fig. 1E) was investigated. As shown in Fig. 1F, the expression level of YAP1 in patients with lymph node metastasis (N1-3) was significantly higher than that in patients without lymph node metastasis (N0). In addition, the GSE database used to analyze YAP1 expression levels in colon cancer patients with or without distant metastasis (Fig. 1G) demonstrated that the expression of YAP1 was significantly enhanced in patients with distant metastasis compared to patients without distant metastasis. Finally, the correlation between YAP1 expression and distant metastasis-free survival (DMFS) in colon cancer patients revealed that the DMFS of patients with high YAP1 expression was significantly lower than that of patients with low YAP1 expression (Fig. 1H). These results demonstrate that altered expression levels of YAP1 affect cancer metastasis and reduce patient survival rates in the development and progression of colon cancer.
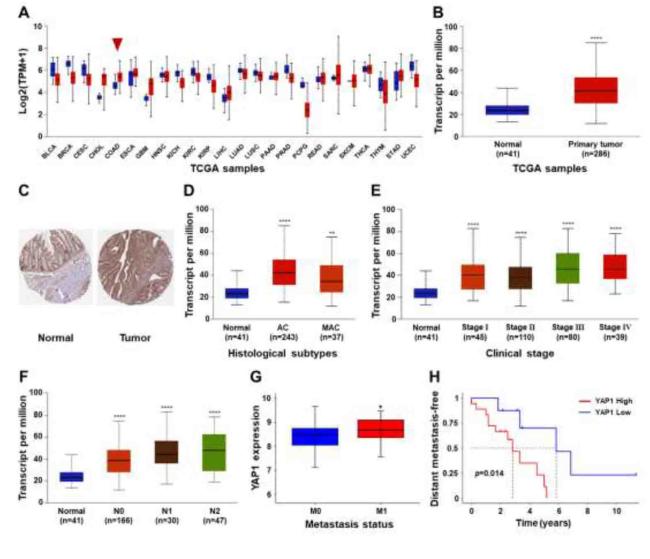
Fig. 1. YAP1 expression was associated with metastasis and poor prognosis in colon cancer (A) Pan-cancer. |
3.2. ECM stiffness enhanced CCCs’ invasion
Studies have shown that ECM stiffness in tumor tissues is closely associated with malignant transformation and cancer.30,31,32 To investigate the effects of ECM stiffness on the migration and invasion capacities of colon cancer cells, we prepared PA hydrogels with adjustable stiffness through oxidation-reduction radical polymerization by changing the concentration ratio of acrylamide and bis-acrylamide, APS initiator, and TEMED catalyst (Fig. S1A). The Young's modulus of the PA hydrogels were measured using a rheometer, as shown in Figure S1 C. A soft hydrogel of 3 kPa was used to simulate normal colon tissue,33 while 20 kPa and 38 kPa hydrogels were used to simulate the ECM stiffness of malignant colon cancer tissues.34 Meanwhile, the cytotoxicities of PA hydrogels to CCCs were detected by the CCK8 method, and the results showed that PA hydrogels were not cytotoxic to CCCs (Figs. S2 and S3), making them suitable for subsequent experiments.
CCCs (HCT116) were cultured into multicellular spheres, which were then inoculated on PA hydrogels of different stiffnesses, and cell adhesion and invasiveness after 4 and 24 h (Fig. 2A and B). After 24 h of sphere culture, the spheres cultured on the 3 kPa soft hydrogel remained spherical with clear edges, a small number of single cells around the spheres invaded the surrounding environment, and the cells in the invaded environment showed a round shape (Fig. 2B). The cell spheres cultured on stiffer hydrogels (20 and 38 kPa) had larger adhesion areas and higher degrees of invasion than those cultured on soft hydrogels (Fig. 2B and C). These data suggest that a stiffer ECM promotes the aggressiveness of CCCs, further proving that ECM stiffness in the tumor microenvironment could be involved in the regulation of tumor cell metastasis. Single-cell tracking analysis showed (Fig. 2E-H) that the motility of CCCs was weakest on the soft hydrogel at 3 kPa. Compared to the cells on the soft hydrogel, the Accumulated distance (Fig. 2F), Euclidean distance (Fig. 2G) and cell movement velocity (Fig. 2H) significantly increased along with enhanced ECM stiffness. Taken together, these results indicate that CCC motility was regulated by ECM stiffness.
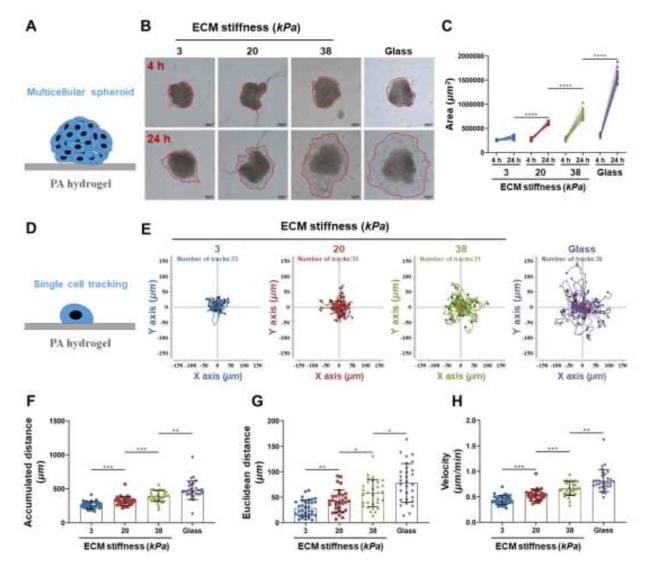
Fig. 2. ECM stiffness increases the invasiveness of cancer cells. |
3.3. ECM stiffness facilitated CCCs' polarization during migration
The first step of cell invasion is the polarization and formation of the front-rear polarity axis, which controls orientation and persistence during cell migration.35 Morphological changes in CCCs cultured on PA hydrogels with different stiffnesses (3, 20, and 38 kPa) at 24, 48, and 72 h were observed using an inverted microscope. As shown in Fig. 3A and B, the cells on the stiffer hydrogels (20 and 38 kPa) gradually became spindle and irregular in shape, and the spreading areas of the cells also increased. In contrast, the volume of cells cultured on the soft hydrogel (3 kPa) decreased, with a low cell spreading area and a round shape. However, cells cultured on the 20 and 38 kPa hydrogels showed no significant difference in cell area between 48 and 72 h. The roundness (Fig. 3C) and aspect ratio (Fig. 3D) of the cells significantly decreased and increased, respectively, along with an enhanced hydrogel stiffness. Thus, ECM stiffness facilitates cell polarization during migration.
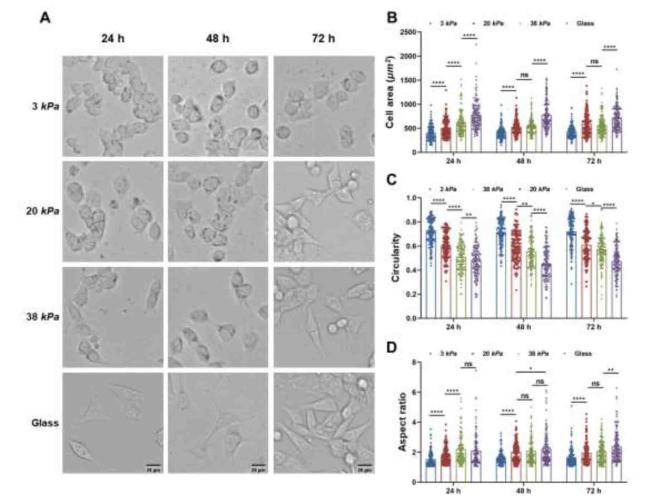
Fig. 3. Time-dependent morphological changes of cell on different ECM stiffness. |
3.4. ECM stiffness tuned mechanophenotypes of CCCs into invasive phenotypes
Mechanophenotypes of cells reflect the relationship between their structure and function. To investigate the effects of ECM stiffness on CCC mechanophenotypes, first, the Young's modulus of cells cultured on hydrogels with different stiffnesses were measured by AFM, as shown in Fig. 4C. The Young's modulus of cells increased with hydrogel stiffness (3, 20, and 38 kPa), but there was no significant difference between the 38 kPa and glass groups (Fig. 4D-G). Second, the EPM reflects the amount of negative charge carried by the plasma membrane. As shown in Fig. 4H, EPM increased significantly along with increase in hydrogel stiffness, indicating that the repulsive forces between the CCCs increased. Third, the membrane fluidity of CCCs was quantified using the fluorescence polarization parameter (p), which was negatively correlated with the motility of membrane lipid molecules. The results showed that the p value decreased with increasing hydrogel stiffness, indicating that the motility of the membrane lipid molecules of the cells significantly increased (Fig. 4I). No significant differences were observed between 3 and 20 kPa. These results demonstrate that the mechanophenotypes of CCCs are regulated by ECM stiffness, which promotes their invasive phenotypes.
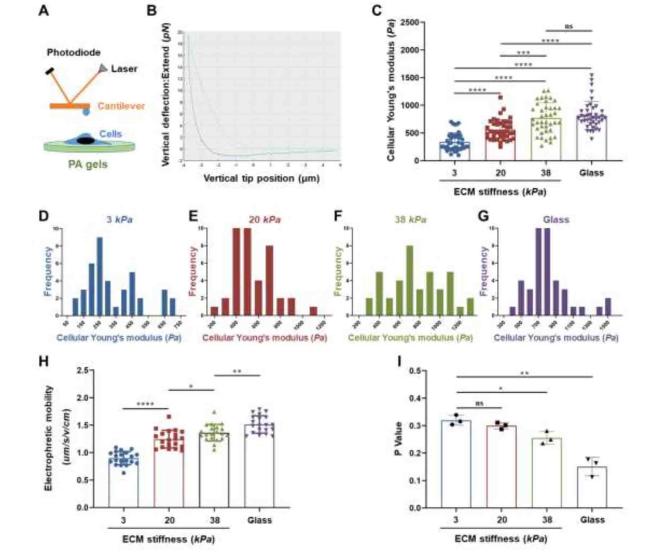
Fig. 4. ECM stiffness affected the cellular biomechanical properties of CCCs. |
3.5. Paxillin clustering in CCCs contributed to F-actin reorganization in stiffness-dependent manner
During cell migration, the dynamic cycle of FA assembly and disassembly at the edge of a migrating cell provides a traction force, the dysfunction of FAs is an important step in tumor cell invasion and metastasis.18,36 Thus, we hypothesized that ECM stiffness promotes CCC migration via abnormal FAs dynamics. First, the expression levels of adhesion-associated proteins (paxillin, talin, and vinculin) and formation of FAs in CCCs cultured on hydrogels of various stiffnesses were investigated using immunofluorescence. The results showed (Fig. S4) that paxillin could be involved in the regulation of FAs formation in CCCs. Therefore, paxillin was used as the FA marker. As shown in Fig. 5A, paxillin was diffusely distributed in the cytoplasm of cells grown on the 3 kPa hydrogel. However, in cells cultured on stiff hydrogels (20 and 38 kPa), paxillin was distributed in dense clusters and accumulated specifically at the ends of fasciculate actin microfilaments. Moreover, CCCs cultured on stiffer hydrogels (Fig. 5A) typically formed polarized stress fiber bundles compared with those cultured on soft hydrogels, and the number of paxillin clusters per cell increased in a hydrogel stiffness-dependent manner (Fig. 5B). The distribution of paxillin in the front-rear polarity axis of the cells was analyzed, and the results (Fig. 5C) showed that paxillin in cells cultured on a soft substrate (3 kPa) was progressively distributed from the edge to the center. With increasing hydrogel stiffness, paxillin is transferred from the center to the edge, leading to FAs formation. In addition, the protein expression levels of paxillin were significantly upregulated in the stiffer hydrogels (Fig. 5D and E). These results indicated that ECM stiffness promotes the formation of FAs at the anterior membrane prominence of CCCs, suggesting that paxillin clusters are involved in cell polarization.
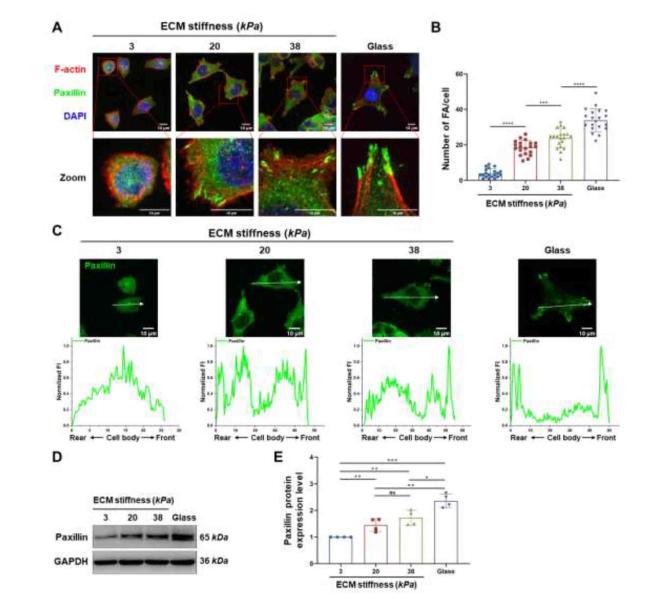
Fig. 5. Stiff ECM promoted focal adhesion formation. |
3.6. ECM stiffness regulated the motility and morphology of CCCs via YAP1 signaling pathway
It is well known that YAP1 is a type of mechanosensitive transcription factor in cells. As shown in Fig. 6A, B, and C, the mRNA and protein expression levels of YAP1 in CCCs cultured on stiffer hydrogels were significantly upregulated. Moreover, the localization of YAP1 was measured by immunofluorescence staining (Fig. 6D) and nuclear-cytoplasmic protein separation by WB (Fig. S5); the results showed that the fluorescence intensities of YAP1 in the cytoplasm of CCCs cultured on soft hydrogels (3 kPa) and stiffer hydrogels (20 and 38 kPa) were significantly stronger and weaker, respectively, than those in the nucleus, while the expression levels of phosphorylated YAP1 (p-YAP1) in the cytoplasm of CCCs cultured on soft hydrogels were significantly higher than those of stiffer hydrogels, indicating that CCCs could respond to changes in ECM stiffness via the YAP1 signaling pathway. To further explore the potential mechanisms of YAP1 in CCCs in response to changes in ECM stiffness, the expression of YAP1 in CCCs was inhibited using the YAP1 inhibitor verteporfin (VP) and short hairpin RNA (shRNA). As shown in Figures S6 and S7, the YAP1 expression in CCCs and cells invasion were effectively inhibited by 1 μM VP and shYAP1-3. Therefore, 1 μM VP and shYAP1-3 were chosen for subsequent experiments. As shown in Fig. 7, multicellular spheroids cultured on stiffer hydrogels (20 and 38 kPa) treated with VP (Fig. 7A and B) and shYAP1 (Fig. 7C and D) showed decreased overall motility (Fig. 7E and F). These results demonstrate that YAP1 is involved in the regulation of CCC morphology and motility.
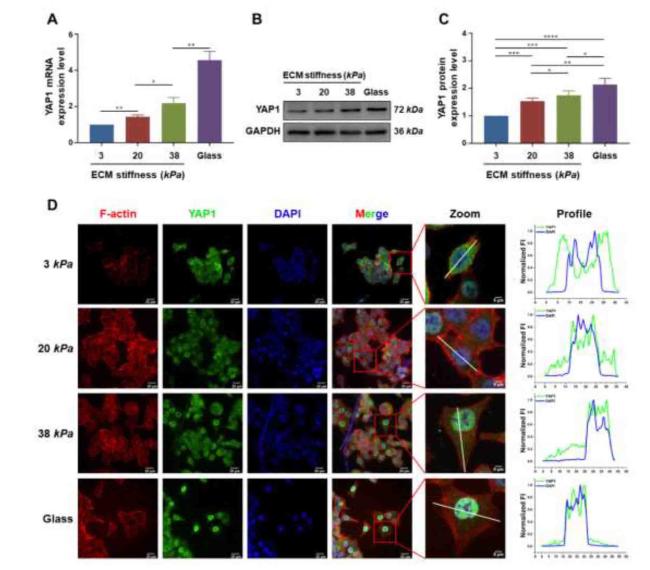
Fig. 6. ECM stiffness promoted the expression and nuclear localization of YAP1. |
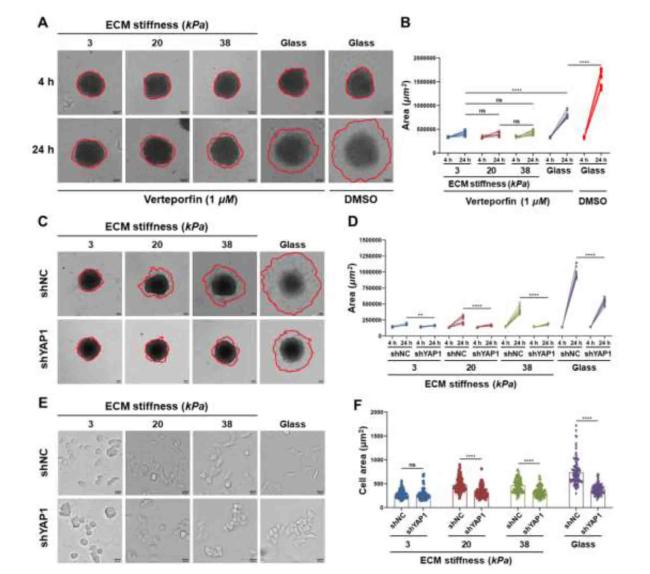
Fig. 7. YAP1 regulated colon cancer metastasis. |
3.7. YAP1 knockdown inhibited FA clustering and altered the mechanophenotypes of CCCs
During cell migration, the assembly and disassembly of FAs play crucial roles in coordination, adhesion, and retraction at the back-trailing edge.37 To investigate the mechanism underlying the function of YAP1 in FAs formation, the correlation between the expression levels of YAP1 and paxillin in colon adenocarcinoma cells was analyzed using the GEPIA database. The results showed a positive correlation between the expression levels of YAP1 and paxillin (Fig. 8B). The immunofluorescence staining results (Fig. 8C) showed that YAP1 knockdown cells significantly reduced the number (Fig. 8D) and size (Fig. 8E) of FAs compared with those of shNC cells, while the higher expression of paxillins was distributed throughout the cytoplasm in shYAP1 cells (Fig. 8F), whereas the lower expression of paxillins along the edge of the nucleus to the cell membrane in shNC cells was recruited for FAs assembly. These results indicated that YAP1 regulates FAs formation at the anterior membrane prominence of CCCs. In addition, YAP1 knockdown led to altered mechanophenotypes of CCCs owing to ECM stiffness, including decreased Young's modulus (Fig. 8G), and membrane fluidity (Fig. 8H).
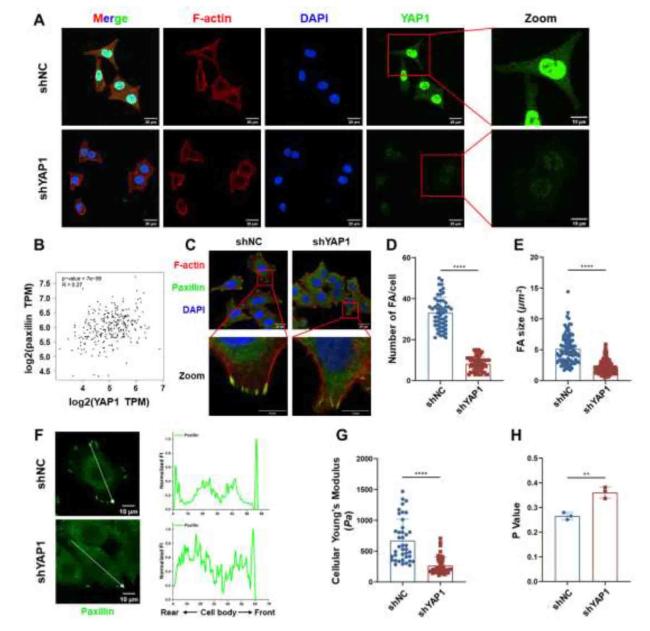
Fig. 8. YAP1 regulated focal adhesion clustering and cellular biomechanical properties of CCCs. |
4. Discussion
Distal metastasis is the primary cause of treatment failure in colon cancer.38,39 Most studies have focused on the chemical factors that regulate the invasion and metastasis of CCCs, including cytokines, the microbiome, and free radicals.40 However, the regulatory mechanism of the impact of changes in the physical properties (e.g., stiffness) of the tumor microenvironment, induced by peroxidation of the extracellular matrix by free radicals generated by the vigorous metabolism of tumor cells, on the invasion and metastasis of CCCs is not fully understood. In this study, bioinformatics analysis showed that the higher expression level of YAP1 in colon cancer was significantly associated with invasion, metastasis, and reduced patient survival rates (Fig. 1). Therefore, it could be hypothesized that YAP1 may serve as a molecular target for the clinical treatment of distant metastasis of colon cancer.
To determine the direction of cell migration, cells first acquire a characteristic polarization pattern in response to extracellular signals.41 As shown in Fig. 2, Fig. 3, the morphologies of CCCs, including the spreading area, circularity, and aspect ratio, were significantly positively correlated with their ECM stiffness, suggesting that CCCs could regulate their shapes in terms of changes in ECM stiffness, which is consistent with previous reports that ECM stiffness affects cell morphology.42,43 Interestingly, CCCs cultured on a stiffer ECM exhibited a highly polarized morphology compared to those cultured on a soft ECM.
The mechanophenotypes (biomechanical characteristics) of cells change with the physical and chemical extracellular microenvironments, reflecting the relationship between their structure and function.44 The Young's modulus of cells is an important mechanical characteristic that reflects their stiffness.45 As shown in Fig. 4D-G, the Young's modulus of CCCs increased with ECM stiffness enhancement, which might be mainly due to the resistance of CCCs to increased ECM-derived mechanical forces to maintain their stable morphology, which is consistent with other recent studies on cervical cancer cells.42 Second, several studies have found that the invasion and metastasis potential of cancer cells is closely related to EPM; the higher the EPM, the stronger the invasion and metastasis capabilities of the cells.46 The EPMs of CCCs were enhanced by a stiffer ECM (Fig. 4H), indicating more negative charges of CCCs and more repulsion force between cells, thus promoting the invasion and metastasis of cancer cells. Third, the fluidity of cell membranes is a necessary condition for maintaining physiological functions such as migration, reflecting the dynamic changes in lipid molecules in the plasma membrane,47 which is associated with the invasion and metastasis of cancer cells.48,49 As shown in Fig. 4I, the membrane fluidity of CCCs increased significantly with increasing matrix stiffness, which is beneficial for the invasion and metastasis of cancer cells. Based on the above results, alterations in ECM stiffness alter the mechanophenotypes of CCCs, resulting in invasive phenotypes.
Dupont et al. studied the relationship between the YAP1 transcription factor and ECM stiffness and found that YAP1 is a sensor of ECM stiffness-derived mechanical force from the cellular microenvironment.28 YAP1 regulates the expression of target genes with a wide range of functional effects in many cell and tissue types. The YAP1 signaling pathway is critical for maintaining cellular functions and normal tissue homeostasis, and an imbalance or failure in this process is involved in the occurrence and development of various diseases.34 Here, polyacrylamide hydrogels with different stiffnesses were used to simulate normal colon tissue and abnormal colon cancer tissue to further investigate the effect of ECM stiffness colon cancer tissue on CCCs’ metastasis via the YAP1 signaling pathway. It has been shown that the colocalization of mechanical sensors at FAs sites enables mechanical and biochemical coupling, leading to sensitivity to changes in extracellular mechanical microenvironments.16 During migration, cells undergo polarization and form protrusions at the front, where new FAs are formed at protrusion sites. FAs assembly plays a critical role in directed cell migration, and the formation of FAs at the leading edge provides traction against the generation of tensional forces that promote cell migration.37 As shown in Fig. 5, Fig. 6, the stiffer ECM promoted the formation and maturation of FAs and upregulated the expression levels of paxillins at the membrane protrusion of cells. Furthermore, as crucial molecules for FAs assembly and disassembly, the paxillins mainly colocalized with FAs at the front protrusion of cells along with the increase in ECM stiffness, inferring that the colocalization of FAs and paxillins at the front protrusion of CCCs could be the upstream signal of YAP1. The expression levels and localization of YAP1 in CCCs cultured on different ECM stiffnesses were measured, as shown in Fig. 7; the expression levels of YAP1 in CCCs were markedly upregulated by stiffer ECM, whose localization translocated from the cytoplasm to the nucleus, indicating that YAP1 worked for the changes in ECM stiffness. After inhibiting and silencing YAP1 (Fig. 8), the CCCs showed reduced migration behavior and altered mechanophenotypes, indicating that YAP1 plays key roles in the invasion and metastasis of CCCs in a stiffer ECM microenvironment. Breast cancer cells also show a similar phenomenon.50 It is possible that YAP1 could serve as a clinical therapeutic target for reducing the invasion and metastatic capability of solid cancer ECM, and anti-fibrotic drugs such as pirfenidone, which downregulate ECM stiffness, may serve as potential clinical treatment options for reducing the distant metastasis of solid cancer cells.
5. Conclusion
In this study, we found that higher expression levels of YAP1 were markedly associated with the invasion and metastasis of CCCs and poor prognosis of colon cancer patients, and that ECM stiffness influenced the invasion and metastasis capability and mechanophenotypes of CCCs via the FA-paxillin-YAP1 signaling pathway. It is important to further understand the invasion and metastasis mechanisms of CCCs, which have laid the theoretical foundation for proposing new clinical protocols for colon cancer from the perspectives of mechanobiology and mechanomedicine.
Author contributions
Kaide Xia: Conceptualization, methodology, data curation, investigation, formal analysis, writing, and Original Draft. Wenhui Hu: Formal analysis, methodology, and writing the Original Draft. Yun Wang: Project administration and resources. Jin Chen: Project administration and resources. Zuquan Hu: Project administration; funding acquisition; supervision; validation. Chenyi An: Methodology and Resources. Pu Xu: Investigation and methodology. Lijing Teng: Investigation, methodology, and resources. Jieheng Wu: Investigation; methodology. Lina Liu: Investigation and resources. Sichao Zhang: Methodology and resources. Jinhua Long: Project administration, supervision, validation, writing, reviewing, and editing. Zhu Zeng: Project administration, conceptualization of resources, supervision, funding acquisition, writing, Review & Editing.
Ethical approval
The CCCs (HCT116) were donated by the Institute of Oncology, Affiliated Tumor Hospital of Guangzhou Medical University (Guangzhou, China). The protocol was approved by the Human and Animal Ethics Committee of Guizhou Medical University (No. 2023-46).
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgement
This study was supported by the National Natural Science Foundation of China (grant numbers 12132006, 32371373, 31771014, and 82060555).
Appendix A. Supplementary data
Supplementary data to this article can be found online at https://doi.org/10.1016/j.mbm.2024.100062.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}