
1. Introduction
Osteocytes, the most abundant cells in bone, are primary mechanosensing cells that regulate bone formation and resorption.1 Due to its network system, bone deformation generated by physical activities creates fluid pressure gradients in its lacunar-canalicular system. The fluid flow generated by these pressure gradients induces shear stress on the osteocyte membrane surface.2 Even though various fluid flow patterns, such as steady fluid flow or oscillatory flow, are happening in the lacunar-canalicular system where osteocytes reside, steady fluid flow as one of the mechanical stimuli has been found to have relatively higher stimulatory to bone cells than other fluid flow patterns in vitro.2,3 The flow pattern in vivo is probably a steady flow superimposed on an oscillatory flow. Analytical modeling study using imaging technique estimated that the peak shear stress on the osteocyte process generated by fluid flow could reach 5 Pa,4 and our previous work found that 4 Pa SFF stimulation was able to generate repetitive intracellular calcium responses.5 Those shear stress-stimulated osteocytes can increase bone formation through anabolic proteins such as sclerostin and decrease bone resorption through catabolic signals such as receptor-activated NF-κB ligands (RANKL) and associated antagonist osteoprotegerin (OPG).6 Our previous study showed that mechanically stimulated osteocytes enhanced the release of extracellular vesicles (EVs) through intracellular calcium oscillations, and these released EVs may facilitate downstream signaling pathways regulation and mechanotransduction between bone cells.7
EVs are nanometer-sized lipid particles released extracellularly, and the role of EVs in bone cell signaling transduction has been studied in recent years.7,8,9 These EVs have been reported to carry proteins, RNAs, and DNAs that function in bone remodeling regulation.10,11,12,13 The studies focusing on EVs released by osteocytes are mainly microRNAs-containing EVs.12,14,15 Still, more recent studies have found that bone-related proteins, such as sclerostin, OPG, and RANKL, have also been detected in EVs released by MLO-Y4 cells in vitro.7 Even though the MLO-Y4 cell line is the most widely used osteocytic cell line in vitro, the relatively low expression levels of Sost and Dmp1, osteocyte mature markers, limit its potential to study the mechanobiology of osteocytes.16 Moreover, the sclerostin, encoded by the Sost gene, is a critical protein for regulating the osteoblastic Wnt/β-catenin signaling pathway for osteogenic activity.17 Lacking the expression of Sost hinders the communication between osteoblasts and osteocytes. A recent study suggested the potential of OCY454 cells for a novel osteocyte mechanobiology study cell model that can respond to oscillatory fluid flow (OFF) stimulation by releasing sclerostin.6 Most importantly, the roles of mechanically induced EVs released by osteocytes in directly regulating osteoblast and osteoclast functions remain undiscovered. Thus, in this study, we are using the OCY454 cell line with high expression of the Sost gene to study the functions of mechanically induced EVs release by osteocytes in regulating osteoblast and osteoclast activity.
2. Materials and methods
2.1. OCY454 cell culture
Osteocyte-like OCY454 cells (gift from Dr. Paola Divieti Pajevic, Boston University, Boston, MA, USA) were grown on 100 mm tissue culture dishes in minimum essential alpha medium (α-MEM, Gibco, Grand Island, NY) supplemented with 10% fetal bovine serum (FBS, Gibco, Grand Island, NY) and 1% antibiotic-antimycotic (Anti-Anti, Gibco, Grand Island, NY). OCY454 cells were seeded at 1 × 105 per 100 mm dish and incubated at 33 °C, 5% CO2 for three days. Then cells were transferred to 37 °C, 5% CO2 for 14 days of maturation without cell passaging.6 Dmp1-GFP expression was checked under the fluorescent microscope before the flow experiment.
2.2. Intracellular calcium response quantification
After 14 days of 37 °C incubation, OCY454 cells were trypsinized and seeded on glass slides at a density of 1 × 106 per slide and incubated overnight. The cells were washed with PBS for 3 times and stained with Fluo-8 AM (ATT Bioquest, Sunnyvale, CA) Ca2+ indicator for 45 min before imaging. Seeded and stained glass slides were inserted into a customized flow chamber and subjected to steady unidirectional fluid flow at 4 Pa shear stress for 9 min. Fluorescent intensity was recorded with a high-speed CCD camera (ORCA-ER-1394, Hamamatsu City, Japan) for 10 min. Baseline intensity was calculated based on 1 min rest period before fluid flow stimulation. Individual cells were selected, the backgrounds were removed, and the average fluorescent intensities were calculated using MetaMorph software (Molecular Devices, Downingtown, PA). The intracellular calcium intensity was normalized by baseline intensity. Responsive cells and the number of peaks were counted as having greater than the mean magnitude of the calcium response (1.2 times fold-change compared to baseline fluctuations) using Matlab.
2.3. Parallel plate flow chamber experiment
OCY454 cells were seeded on glass slides at 1 × 106 cells per slide and incubated at 37 °C overnight. Before fluid stimulation, cell-seeded slides were rinsed three times with PBS. Six cell-seeded slides were inserted into a custom-made flow chamber for steady fluid flow (SFF) stimulation with 4 Pa shear stress in α-MEM supplemented with exosome-depleted FBS (Gibco). Cells underwent one (1C), two (2C), and three (3C) 10-min bouts of steady fluid flow separated by one 15-min rest period in 2C and two 15-min rest periods in 3C (Fig. 2B). Before the 10-min bout started, a 15-min rest period was used to stabilize cells. Rest insertions are mechanically stimulating for bone formation in vivo.18,19 Control slides were sealed in dishes with an equal volume of the fluid medium during the fluid flow experiments. After 75 min of processing time, cells and supernatant from all the groups were collected immediately for gene expression, protein expression and co-culture experiments.
2.4. Isolation and characterization of extracellular vesicles
The supernatant collected after fluid flow stimulation experiments was served as the conditioned medium and frozen at −20 °C before analysis. EVs were isolated from 48 mL conditioned medium by differential ultracentrifugation20 as previously described.7 Isolated EVs were resuspended in 1 mL PBS for EVs characterization or 48 mL fresh flow medium served as the EVs-only medium. The supernatant collected after the last step of ultracentrifugation was used as the EVs-free culture medium in the following osteoblast and osteoclast co-culture experiments. Nanosight (Malvern, United Kingdom) has been used to measure the size and concentration of extracellular vesicles. EVs contents were assessed by enzyme-linked immunosorbent assay (ELISA). Protein lysate of EVs was prepared by resuspending EVs pellet in RIPA buffer for 15 min. The concentration of sclerostin, RANKL, and OPG in the medium was measured by sclerostin ELISA kit (R&D System), RANKL ELISA kit (Invitrogen), and OPG ELISA kit (Invitrogen). The final protein concentration in EVs lysate was adjusted to the volume of the medium it isolated from.
2.5. Osteoblast conditional experiment and quantification of extracellular ALP activity
Osteoblast MC3T3-E1 cells were maintained at 37 °C in a humidified 5% CO2 atmosphere in a growth medium (alpha-MEM, 10% FBS, and 1% Penicillin-streptomycin) and passaged at less than 80% confluence. Before the osteoblast conditional experiment, MC3T3-E1 cells were seeded at 5 × 103 per well in a 24-well plate and cultured under 50% osteogenic medium and 50% conditional medium (full conditioned medium or EVs-free medium or EVs-only medium). The same formulation replaced the culture medium on days 4, 7, and 10. The osteogenic medium was a growth medium supplemented with 50 μg/mL ascorbic acid (Sigma-Aldrich) and 10 mM ꞵ-glycerophosphate (Sigma-Aldrich). On day 10, supernatants were collected, and extracellular ALP activity was quantified using an alkaline phosphatase activity colorimetric assay kit (APExBIO) according to the manufacturer's recommendation. Briefly, 5 mM p-nitrophenyl phosphate (pNPP) was added to samples and incubated for 1 h. Synergy 4 detection system (BioTek, USA) was used to measure absorbance at 405 nm.
2.6. Osteoclast conditional experiment and tartrate-resistant acid phosphatase (TRAP) staining
RAW264.7 cells were maintained in a growth medium (D-MEM/High glucose with l-glutamine (Cytiva) containing 10% fetal bovine serum and 1% penicillin-streptomycin (PS, Gibco, USA) at the atmosphere of 5% CO2 and 37 °C. Cells were passaged by scraping cells vigorously after reaching 80% confluency. Before the osteoclast conditional experiment, RAW264.7 cells were seeded at 2.5 × 103 per well in a 24-well plate and cultured under 50% osteoclast differentiation medium (growth medium supplemented with 20 ng/mL recombinant RANKL (R&D Systems) and 50% conditional medium (full conditioned medium or EVs-free medium or EVs-only medium). The same formulation replaced the culture medium on days 4, 5, and 6. Osteoclasts were visualized using TRAP staining on day 7. Briefly, cells were fixed with 4% paraformaldehyde on day 7 and stained with the leukocyte acid phosphatase kit (Sigma-Aldrich, USA). TRAP-positive cells with more than five nuclei were counted as mature osteoclasts.
2.7. Quantitative real-time PCR
After flow stimulation, 3 stimulated slides were used to isolate total RNA by RNeasy Mini Kit (Qiagen, USA), and cDNA was obtained by reverse-transcribed by High Capacity cDNA Reverse Transcription Kit (Applied Biosystems). PowerUp SYBR Green Master Mix (Applied Biosystems) was used to quantify gene expression levels. Primer sequences are available upon request. β-actin was used for the normalization of gene expression. ΔCT was computed within each sample to the housekeeping reference, and ΔΔCT was computed across experimental conditions. Experiments were run in triplicates unless otherwise indicated.
2.8. Western blot
OCY454 cells, after fluid flow stimulation, were lysed in RIPA buffer supplemented with a protease/phosphatase cocktail (ThermoScientific, Waltham, MA). Protein concentrations were quantified by Pierce BCA Protein Assay Kit (ThermoScientific, Waltham, MA), and 18 μg protein was separated on a 12% Mini-PROTEAN TGX Precast Protein Gels (Bio-Rad, USA) and transferred to a PVDF membrane according to the manufacturer's recommendations. The membrane was blocked with 5% nonfat milk (Bio-Rad) in Tris-buffered saline containing 0.05% Tween 20 (TBST) for 1 h. Primary antibodies, anti-sclerostin (Invitrogen), anti-OPG (Invitrogen), anti-RANKL (R&D System), and anti-GAPDH (Invitrogen), were incubated with membrane overnight at 4 °C. After washing, horseradish peroxidase-conjugated secondary antibodies were incubated for 1 h at room temperature and then developed using enhanced SuperSignal West Dura Extended Duration Substrate (ThermoScientific, Rockford, IL) and LAS 4000 imaging system (Fujifilm, USA). The image was processed using Image J (NIH, USA).
2.9. Sclerostin, OPG, and RANKL enzyme-linked immunosorbent assay (ELISA)
Sclerostin (Mouse/Rat SOST Quantikine ELISA, R&D System, USA), OPG (OPG (TNFRSF11B) Mouse ELISA Kit, Invitrogen, USA), and RANKL (TRANCE (RANKL/TNFSF11) Mouse ELISA Kit, Invitrogen, USA) were quantified using ELISA kits on conditioned medium and EVs lysates collected after osteocytes exposed to 4 Pa fluid flow simulation according to manufacturer's recommendation. Briefly, protein was added and incubated for 2 or 3 h, then incubated with HRP-conjugated secondary antibodies for 1 h. TMB substrate was added, and a Synergy 4 detection system (BioTek, USA) was used to measure absorbance at 450 nm.
2.10. Statistical analysis
All values were reported as the mean ± S.D. The significance between the control and each treatment was evaluated with one-way ANOVA followed by Dunnett's multiple comparison test. Statistical significance was considered at P < 0.05. GraphPad PRISM was used for statistical analysis. At least three individual experiment trials were run.
3. Results
3.1. Intracellular calcium response after unidirectional fluid flow stimulation
As we know, mechanically induced calcium response has been verified in both ex vivo and in vivo,7,21 and this response has been used as an indicator of the early mechanotransduction cascade.6 Previous studies have verified that 4 Pa fluid flow stimulation could generate enough fluid shear stress to induce distinguishable intracellular calcium response in osteocytes.5,6 Our previous work found that steady fluid flow has a higher stimulatory effect than oscillatory fluid flow.2 Thus, for the new osteocytic cell line OCY454, we examined the intracellular calcium response under 4 Pa SFF stimulation (Fig. 1A). Calcium oscillation was tracked under a fluorescence microscope, and positive cells and the number of peaks were counted. We found OCY454 cells have intracellular calcium oscillation in response to 4 Pa SFF stimulation (Fig. 1B and C) with 80% responsive cells averaging 3 peaks in a stimulation period of 9 min (Fig. 1D). This suggests that OCY454 cells, similar to MLO-Y4 cell line,22 can respond mechanical stimulation through calcium oscillation.
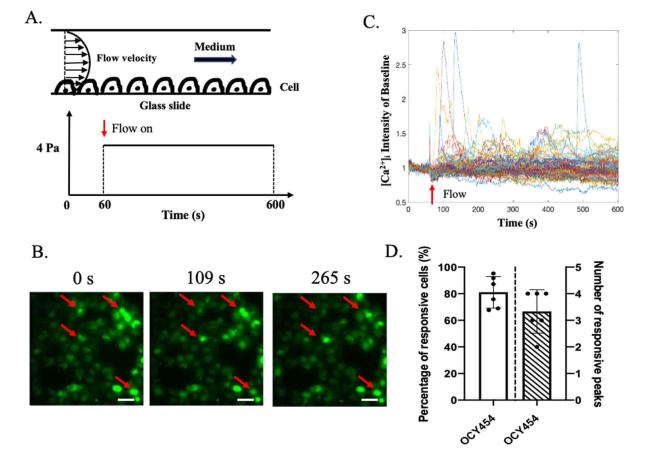
Fig. 1. Intracellular calcium response. 4 Pa unidirectional fluid flow stimulation was applied to OCY454 cells and calcium response was measured simultaneously. (A) OCY454 cells were seeded on glass slide and stimulated with laminar fluid flow. Camera was on through 0 to 600s, and flow was from 60 to 600s. (B) Representative images of intracellular calcium intensity at 0s, 109s and 265s. Red arrows indicate responsive cells. (C) Time course of Fluo-8 AM intensity normalized with baseline. (D) Percentage of responsive cells and number of responsive peaks. n = 6 slides/group. Data were presented as the mean ± SD. Scale bar: 20 μm. |
3.2. Sclerostin, OPG, and RANKL expression in OCY454 cells after fluid flow stimulation
To study the response of OCY454 cells to mechanical loading in different loading durations, one-cycle burst (1C), two-cycle bursts (2C), and three-cycle bursts (3C) stimulation patterns have been used to apply SFF on OCY454 cells (Fig. 2A and B). After 75 min of processing, cells, and supernatant from all the groups were collected immediately. From qPCR results, we found that Sost gene expressions have been downregulated, and Opg gene expression has been upregulated in stimulated OCY454 cells compared with the non-stimulated control group (Fig. 2D). Moreover, this effect was does-dependent on the loading duration. Unlike Sost gene expression, which decreased in all the stimulation groups, Rankl gene expression has been upregulated at the beginning and downregulated afterward in the 2C and 3C groups. The Opg/Rankl ratio has increased significantly after stimulation. Even though Dmp1, one of the mature osteocyte marker genes,23 mRNA expression level has increased trend, they did not have statistical significance in all the groups (Fig. 2C). Similar to mRNA expression results, sclerostin protein levels in cells were decreased in flow stimulated 3C group and the concentration of sclerostin in the medium was reduced in flow stimulated group (undetectable) compared with the control group (Fig. 3A, B, and C). OPG levels in cells and concentration in the conditioned medium were upregulated in response to mechanical stimulation (Fig. 3A, B, and C). OPG/RANKL ratio in cells has significantly elevated in response to 2C and 3C fluid flow stimulation (Fig. 3B). However, no RANKL has been detected in the conditioned medium (Fig. 3C). These results suggest that sclerostin and OPG/RANKL ratio have been regulated in response to SFF stimulation in OCY454, and this regulation was load-duration dependent.
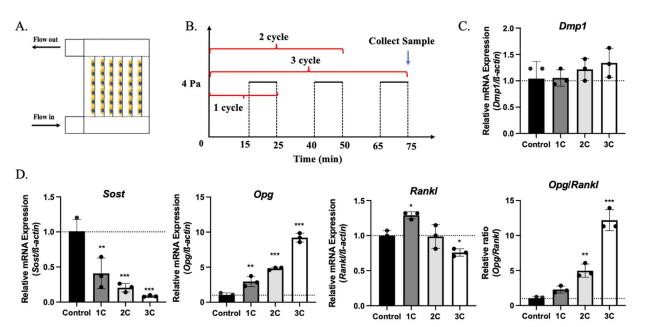
Fig. 2. Gene expression in OCY454 after fluid flow stimulation. (A) Schematic of the multi-slide fluid flow chamber. (B) Fluid flow profile of control, 1 cycle (1C), 2 cycle (2C) and 3 cycle (3C) groups. Gene expression of Dmp1 (C), Sost, Opg and Rankl (D) in OCY454 cells after 1C, 2C, and 3C 4 Pa fluid flow stimulation. n = 3 samples/group. ∗p < 0.05, ∗∗p < 0.01, ∗∗∗p < 0.001. Data were presented as the mean ± SD. |
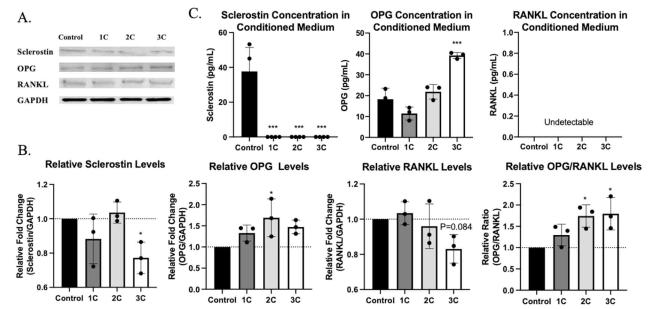
Fig. 3. Protein expression in OCY454 cells and in the conditioned medium after 4 Pa fluid flow stimulation. (A) Western blots for sclerostin, OPG, RANKL and GAPDH proteins in OCY454 cells after mechanical stimulation. (B) Quantification of protein levels from western blots. (C) Sclerostin, OPG and RANKL concentration in conditioned medium after fluid flow stimulation. n = 3 samples/group. ∗p < 0.05, ∗∗∗p < 0.001. Data were presented as the mean ± SD. |
3.3. Extracellular vesicles release and protein expression in stimulated OCY454 cells
Our previous study found that mechanically induced calcium oscillation enhanced extracellular vesicles (EVs) release in MLO-Y4 cells.7 Thus, we also characterized the EVs released from OCY454 by ultracentrifuging the conditioned medium.20 Mechanical stimulation significantly increased EVs release into the conditioned medium, and the concentration of EVs is dose-dependent on the loading duration (Fig. 4C). The average size of released EVs was around 120 nm (Fig. 4A and B). This indicates that those released EVs were exosomes or microvesicles.24 To understand the role of EVs in osteocytic mechanical regulation, EVs pellets were lysed by resuspended in RIPA buffer, and protein levels in EVs have been measured by ELISA kit. Interestingly, opposite with conditioned medium, sclerostin was undetectable in EVs lysate, but RANKL was detected and decreased in stimulated groups. Even though a few OPG were detected in stimulated groups, the concentration of OPG was low and negligible (Fig. 4D). This may suggest that osteocytes released RANKL, not OPG or sclerostin immediately in response to flow stimulation through EVs capsulation.
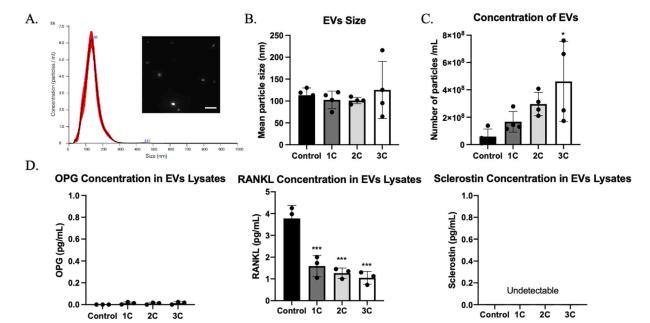
Fig. 4. Extracellular vesicles (EVs) characterization. (A) Representative distribution of particle size. (B) Average particle size for each group (n = 4 samples/group). (C) EVs concentration in each group (n = 4 samples/group). (D) OPG, RANKL, and sclerostin concentration in EVs lysates (n = 3 samples/group). ∗p < 0.05, ∗∗∗p < 0.001. Data were presented as the mean ± SD. Scale bar: 400 nm. |
3.4. Stimulated OCY454 cells regulate osteoblast and osteoclast functions
Osteocytes, as major mechanical sensing cells in bone, transform mechanical stimulation into cell signaling to regulate osteoblast and osteoclast functions and, therefore, maintain bone homeostasis.25 Thus, in order to study the cross-talk between osteocytes and other bone cells, conditioned medium, EVs-free medium, and EVs-only medium from the supernatant of stimulated OCY454 cells were supplemented into MC3T3-E1 and RAW264.7 differentiation medium. Alkaline phosphatase (ALP) is the enzyme secreted by osteoblasts and is known as an early osteogenic marker of bone calcification and mineralization.26 ALP activity in co-culture medium and the number of TRAP+ multi-nuclei cells have been measured to study the effects on osteoblast and osteoclast functions. After 10 days of culture, ALP activity was upregulated in the stimulated conditioned medium and EVs-free medium group but not in the EVs-only group (Fig. 5). In the osteoclast differentiation experiment, decreased TRAP+ multi-nuclei cells were found in flow stimulation group, compared with control which has more multi-nuclei TRAP+ cells (Fig. 6A, B, and D). However, in the groups treated with EVs-free medium, there was no significance between groups (Fig. 6C), suggesting that osteocytes regulate osteoclast differentiation in response to mechanical loading mainly through EVs form, and regulate osteoblast activity through soluble free protein.
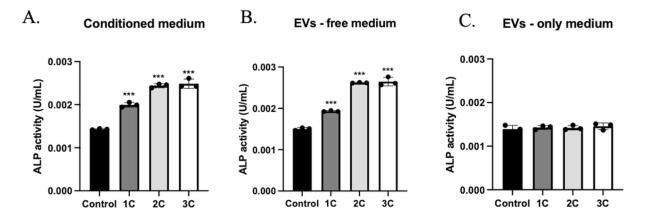
Fig. 5. The effects of conditioned medium with or without EVs and EVs only in osteoblast activity. ALP activity in MC3T3-E1 cells treated with conditioned medium (A), conditioned medium without EVs (B) and EVs only medium (C) for 10 days. n = 3 samples/group. ∗∗∗p < 0.001. Data were presented as the mean ± SD. |
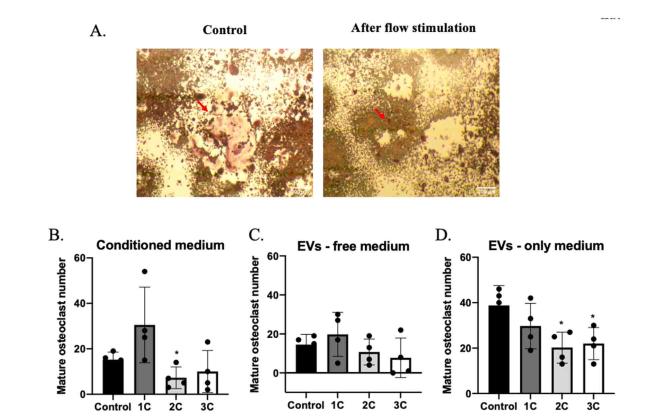
Fig. 6. The effects of conditioned medium with or without EVs and EVs only in osteoclast maturation. (A) TRAP stain in RAW263.7 cells treated with fluid flow medium (right) or static medium (left). Mature osteoclast numbers (nuclei ≥5) in conditioned medium (B), conditioned medium without EVs (C), and EVs only medium treated groups (D). n = 4 samples/group. ∗p < 0.05. Data were presented as the mean ± SD. Scale bar: 200 μm. |
4. Discussion
In this study, we used a new osteocytic cell line, OCY454, to characterize the function of mechanically induced EVs release of osteocytes in osteoblast and osteoclast regulation. This new cell line has been reported with high sclerostin expression, and using this cell line provided the opportunity to study the osteocytes-osteoblasts and osteocytes-osteoclasts communications in vitro.6 However, as the mechanosensor embedded in the bone matrix, the mechanism of osteocytes translating mechanical changes in bone and further regulating bone adaption through osteoblast and osteoclast is still unclear. Our previous work suggested that mechanically induced intracellular calcium oscillation in osteocytes facilitates EVs release and regulates bone metabolism by the protein release through EVs.7 Thus, we examined the role of those released EVs by stimulated OCY454 cells in regulating osteoblast and osteoclast functions.
Here, we focused on instantly released EVs regulating osteoblast and osteoclast functions. After 4 Pa fluid flow shear stress stimulation, OCY454 cells enhanced osteogenesis activity by secreting less sclerostin, a negative regulator of osteoblast activity through Wnt/β-catenin signaling pathway, to the extracellular matrix (conditioned medium). These secreted sclerostin were not detected in EVs lysates, suggesting that OCY454 cells instantly regulate osteoblast activity through soluble free sclerostin protein, not EVs capsulated sclerostin. Moreover, ALP as an early osteogenic marker of bone calcification secreted by osteoblast, its activity has no difference between stimulation groups and control groups in EVs-only treated osteoblast medium. This may suggest that instantly released EVs are not essential for osteocyte-regulated osteogenesis activity. Furthermore, in the osteoclast co-culture experiment, the conditioned medium and EVs-only medium from stimulated groups inhibited mature osteoclast numbers, but the EVs-free medium did not. This suggests the importance of EVs in osteocyte-regulated osteoclastogenesis.
EVs are membrane-restricted vesicles released by cells and contain cellular components from parental cells. EVs membranes are formed by lipid bilayers and are able to protect encapsulated contents from degradation through proteases and nucleases.24 Moreover, due to the small size of EVs, they are ideal carriers for transportation. Osteocytes, as the cells embedded in the bone matrix, cannot contact other cells directly for cellular communication. Thus, RANKL, a critical protein for osteoclast maturation, encapsulated by EVs provides a longer circulation time and higher transportation efficiency. This may also explain why EVs-only medium has more obvious osteoclast regulation effects than EVs-free medium, even though fluid flow stimulation increased OPG concentration in conditioned medium (Fig. 3C) and protein concentration is much lower in EVs lysate (Fig. 4D and Fig. S2). Interestingly, gene expression of Rankl was increased first in the 1C group, then decreased in the 2C and 3C groups (Fig. 2D). This increased Rankl expression in 1C may facilitate mono-nuclei TRAP+ cell formation, as a recent study found that mechanical loading induced elevation of mono-nuclei TRAP+ cell number to enhance osteogenesis activity through TGF-β pathway.27 However, these increased RANKL levels were not detected in EVs, which may suggest those increased RANKL were not in EVs form. Thus, RANKL in different forms may have different functions in bone regulation.
While our study shows new findings of the role of EVs released by stimulated OCY454 cells in osteoblast and osteoclast function, there are some limitations, and further work may be necessary. Even though we found significant changes in gene expression in all the stimulated groups, intracellular protein expression in Western blot results did not. This may be due to the short stimulation time to have significant translational changes or the differences in detection sensitivity between methods. However, cell protein expressions still have a similar trend to gene expression. Moreover, these gene expression changes have also been verified in the protein expression after overnight culture (Fig. S1). Furthermore, not only the proteins we have measured in this study, other proteins such as cathepsin K,28 DKK1,29 TGF-β and BMPs30 have all been reported to play a role in bone mechanical responses. We did not exclude the roles of those proteins involved in our results, even though they may have limited functions in the study. This may explain our osteoblast co-culture results that there is a similar ALP activity baseline in the EVs-only medium group compared with the EVs-free medium group. We found that the conditioned medium in the 2C group suppressed osteoclast maturation but not in the 3C group (Fig. 6B). This may be due to other factors released in the 3C group to control osteoclast maturation. Long-term loading in the 3C group may induce osteocyte apoptosis and recent studies reported that osteocyte necrosis would enhance the release of damage-associated molecular patterns (DAMPS) and promote osteoclastic activities by RANKL-independent pathways.31,32 Since EVs are characterized as cellular signal carriers, they may have the potential to induce osteoprogenitors’ migration to the active bone surface. Thus, migration factors such as IGF-133 may also participate in the function of released EVs.
In summary, this work characterized the role of EVs released by mechanically stimulated OCY454 cells in regulating osteoblast and osteoclast functions. We discovered that OCY454 cells regulate osteoclastogenesis by releasing EVs and regulate osteogenesis by releasing free-form proteins in response to mechanical stimulation. We found that RANKL released by osteocytes in response to mechanical stimulation was EVs encapsulated, and sclerostin and OPG were released mainly in free forms.
Funding sources
The NIAMS of NIH supports this work through R01 AR069148 and R01 AR062177.
Disclosures
The authors have nothing to disclose.
Ethical approval
This study does not contain any studies with human or animal subjects performed by any of the authors.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgments
Appendix A. Supplementary data
Supplementary data to this article can be found online at https://doi.org/10.1016/j.mbm.2024.100065.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}