
1. Introduction
Dental-maxillofacial malformations have become serious problems in current society, because of their high incidence, and adverse effects on appearance and oral function [1]. Orthodontics, a primary intervention for malocclusion, involves the application of external forces to reposition teeth [2]. The biological foundation of orthodontic tooth movement (OTM) is predicated on force-induced modification of periodontal tissue, encompassing the periodontal ligament (PDL) and alveolar bone. Therefore, elucidating the biological mechanisms underlying periodontal remodeling under mechanical force is critical.
Two counteractive tissue alterations are involved in the tissue remodeling during OTM: (1) the resorption of superfluous alveolar bone on the stress side, coupled with compression of the periodontium, and (2) the renewal and deposition of stromal bone, coupled with stretching of the periodontium on the tension side [3,4]. Mesenchymal stem cells (MSCs) have self-replication and multi-directional differentiation capacity, and consequently can differentiate into a diverse range of tissue cells, including adipose, bone, nerve, and endothelial cells, under specific induction conditions [5,6]. In addition, they can sense external mechanical forces and translate them into biological signals within cells, thereby further inducing differentiation into mature cells with different functions. In dentistry, the mechanism through which force-induced MSCs remodel periodontal tissues has garnered substantial research attention [7,8,9]. However, analysis of the regression of MSCs under mechanical stimulation and the mechanism of periodontal tissue remodeling remains challenging because of differences between the in vivo and ex vivo environments, the intricacy of the biological microenvironment, and the dynamic changes occurring during growth and development [10,11].
Lineage tracing is a crucial tool in developmental biology that enables tissue stem cell properties to be studied. It provides dynamic profiles of specific stem cell progeny and can be used to assess cell differentiation and migration potential [12]. Current genealogical tracing techniques mainly include direct observation, labeling of cells with dyes and radioactive tracers, introduction of genetic markers by transfection or viral transduction, transplantation of cells and tissues, genetic mosaics, cell marking by genetic recombination and lineage tracing with multicolor reporters [13]. The identification of stem cell biomarkers has enabled the use of tissue-specific gene promoter and site-specific recombinase technologies for genetic lineage tracing of specific stem cell populations. Among which, the Cre-loxP system is the most widely used site-specific recombinase system. The mechanism can be classified into constitutive and inducible recombination depending on whether the timing is controlled [14]. The CreERT2-loxP recombinase system can be induced at specific time points to observe the proliferation, differentiation, and migration of labeled cells. Thus, genetic lineage tracing is a reliable method for identifying heterogeneity in stem cell fate decisions in vivo [15]. Gli1+ cells, identified in Gli1-CreERT2; Ai14 mice, have been shown to be important in stem cell involvement in wound healing in extracted teeth [16]. Osterix-expressing cells have been found to label bone progenitor cells at different developmental stages [17].
Collagen proteins are major components of the extracellular matrix. They share a common structural feature of the presence of at least one triple-helix structural domain composed of three intertwined polypeptide chains. These proteins are abundant in the extracellular matrix and play important roles in cell membrane structure and function. Collagens not only shape tissues but also play crucial roles in cell attachment, proliferation, and migration processes [18]. To date, 28 types of collagen with diverse structures, localization, and biological functions have been identified. For example, type I collagen is present in most connective and embryonic tissues [19], and is composed of two chains of collagen type I alpha 1 (COL1A1) and one chain of collagen type I alpha 2 (COL1A2) [20]. This protein is involved in physiological processes, such as bone mineralization and hemostasis [21]. The mutation of COL1 gene affects the rate of type I collagen N-propeptide cleavage, thus interfering with normal collagen fibrillogenesis, resulting in connective tissue diseases and bone diseases, such as Ehrles-Danlos syndrome and osteogenesis imperfecta [22].
In this study, we used an inducible genetic recombination system together with an OTM model to investigate the characteristics of Col1+ cells in adult mice. Our results indicated that these cells do not undergo substantial proliferation under normal conditions, but their numbers increase in response to mechanical regulation, to a degree proportional to the duration of the applied force. Moreover, we used immunofluorescence staining and other techniques to observe the dynamic distribution of the Col1+ cell lineage under orthodontic force. The results indicated the differentiation potential of osteogenic and fibrogenic lineages of Col1+ cells. In addition, a new experimental model and method were created through combining cell lineage tracing technology with an in vivo OTM mouse model, thus supporting future studies on the function of Col1+ cells in tooth movement.
2. Materials and methods
2.1. Animals
This study was conducted on Col1-CreERT2 mice and ROSA26-loxP-stop-loxP-tdTomato mice (6-week-old male C57BL/6). Col1-CreERT2 mice and CG-Gt(ROSA)26Sortm14(CAG-tdTomato)Hze/J mice were gifts from Bin Zhou, Shanghai Institutes for Biological Sciences, Chinese Academy of Sciences [23]. Col1-CreERT2 mice were crossed with tdTomato mice to generate Col1-CreERT2; tdTomato mice. All mice were maintained on a C57BL/6 genetic background. All animals were housed under standard laboratory environmental conditions (room temperature 22-26 °C; humidity 50-55 %). Standardized laboratory pellet food and water were available ad libitum. All experimental protocols were approved by the Ethics Committee of Shanghai Ninth People's Hospital, School of Medicine, Shanghai Jiao Tong University (approval number: SH9H-2023-A92-1).
2.2. Experimental tooth movement and micro-computed tomography analysis
The OTM model was generated as previously described [24]. After mice were anesthetized, orthodontic forces were applied to the maxillary left first molar with a closed coil spring (0.25 mm wire size, 0.76 mm diameter; Innovative Materials & Devices, Shanghai, China) with a force gauge (Cathaysian Dentistry, Zhejiang, China) measuring 10 gravity. The other end of the spring was attached to the mesial incisor via a 0.1 mm wire supported by a self-curing restorative resin (3M ESPE, St. Paul, MN, USA) for reinforcement. The orthodontic appliances were examined daily, and any mice with displacement were excluded from analysis. The animals were euthanized on day 3 and day 7 after OTM. The maxillae were extracted for histological examination. The OTM distance between the midpoint of the M2 mid-marginal ridge and the midpoint of the M1 distal ridge was measured in ImageJ. Maxillary dissections from day 3 and day 7 mouse OTM models were preserved in ethanol and subsequently scanned with a micro-computed tomography scanner.
2.3. Histological staining
Maxillary bone specimens were fixed with 4 % paraformaldehyde for 48 h and demineralized with 10 % EDTA for 5 weeks. After processing, 4 μm continuous paraffin sections were obtained in cross-sections. The alveolar bone of M1 was defined as the region of interest. To analyze the histological changes in the maxilla, we performed anti-tartrate acid phosphatase staining with an acid phosphatase leukocyte kit (Sigma, St. Louis, MO, USA),H&E staining (Beyotime, C0105M) and immunohistochemical staining.
2.4. Immunofluorescence
After deaffinity rehydration, sections were blocked with 3 % H2O2, and antigen recovery was performed in 0.2 % Proteinase K. Sections were pretreated with 10 % bovine serum albumin to block nonspecific binding for 1 h, then incubated with primary antibodies (rabbit anti-Sp7 (Abcam), rabbit anti-Postn (Abcam), and rat anti-Endomucin (Santa Cruz)). After incubation, fluorescent secondary antibodies were applied. For analysis of histological features, sections were examined and photographed under a light microscope (Olympus Corporation, BX-51, Tokyo, Japan) with a digital camera, and positive cells were counted in regions of interest.
2.5. Statistical analyses
All statistical analyses were performed in Prism software (GraphPad). Results are expressed as mean ± standard deviation or mean ± standard error. Quantitative data were assessed with unpaired Student's t-test or one-way ANOVA. P-values <0.05 were considered significant. No statistical methods were used to predetermine sample sizes. Sample sizes were determined on the basis of prior literature and experience with the model system, to ensure accurate assessment of potentially biologically important differences.
3. Results
3.1. Characterization of periodontal tissue remodeling during OTM
To investigate the dynamic distribution and differentiation potential of Col1-expressing cells during OTM, we created Col1-CreERT2/ROSA26-loxP-stop-loxP-tdTomato (Col1-CreERT2; tdTomato) reporter mice. In these mice, Cre expression could be induced in Col1+ cells at various developmental stages through the administration of tamoxifen (Fig. 1A). To achieve the desired effects, we used a previously described induction strategy [24]. Tamoxifen was intraperitoneally injected into 6-week-old control mice and Col1-CreERT2; tdTomato mice. The OTM model was initiated after a 7-day interval. The intact maxillary alveolar bone and tooth tissue were extracted for further analysis on days 0, 3, and 7 after euthanasia (Fig. 1B and C). Both stereoscopic and micro computed tomography images revealed that the molars of mice that did not undergo orthodontic forces were precisely aligned and tightly interlocked. A small gap, measuring approximately 30-50 μm, between the maxillary first and second molars on day 3, was visible to the naked eye. On day 7, a gap covered by soft tissue was observed between the first and second molars of the mice. The distance between the molars was approximately 100-120 μm on day 7, thus indicating the successful construction of OTM model (Fig. 1D-F). The mechanical force led to changes in both PDLs and alveolar bone tissue surrounding the maxillary first molars. We prepared paraffin sections from both OTM and untreated sides of the same mice on day 7 for HE staining, to obtain a comprehensive and accurate picture of the overall structure of periodontal tissues after mechanical force (Fig. 1G-L). Examination of HE stained tissue sections indicated well-arranged alveolar bone tissue around the three roots of the maxillary first molar on the untreated side of the mice (Fig. 1G). Furthermore, vertical sections of the roots revealed a dense and fibrous ring of PDL surrounding each root (Fig. 1H and I). In contrast, increased formation of bone matrix between the three roots of the first molar was observed on day 7 of OTM. The newly formed bone tissues were disorganized, and the presence of many new blood vessels indicated a substantial change in the tissue structure (Fig. 1J-L). Meanwhile, width of the mesial PDL narrowed, and the fibers were compressed, whereas the distal PDL was stretched and widened (Fig. 1K,L). In brief, we successfully established an OTM model for controlling the interplay between osteogenesis and osteolysis by using Col1-CreERT2; loxp-tdTomato mice, which showed notable alterations in the periodontal ligament.
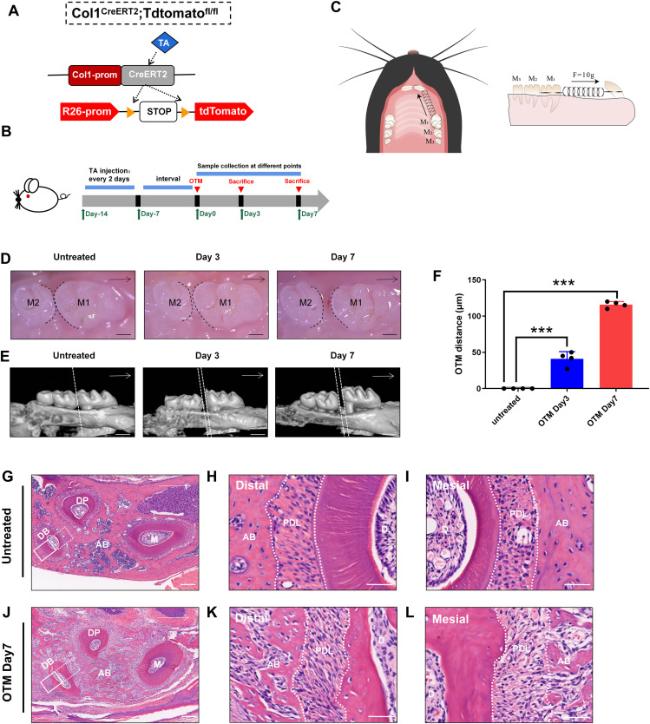
Fig. 1. Mechanical force induces modification of periodontal tissue during OTM. (A) The schematic diagram of the knock-in Col1-CreERT2 mouse induced by tamoxifen (TA). (B) Experimental protocol:Male wild-type (WT) and Col1-CreERT2; tdTomato mice, 6 weeks old, were injected with TA every 2 days (100 mg/kg/bw) starting one week before OTM model construction. Mice were euthanized and specimens were collected on day 3 and 7 after OTM for analysis. (C) The schematic diagram of the orthodontic tooth movement (OTM) device in vivo. The spring ligated between the first molar (M1) and incisors induced orthodontic force of about 10g. (D, E) Images from stereoscope and micro computed tomography showed the distance between M1 and the second molar (M2). (F) The distance between M1 and M2 on the untreated side, d3 and d7 after OTM, which was defined as OTM distance. (G, J) Observation of H-E staining of the alveolar bone of the Untreated side and OTM side of mice on day 7. DB,distal buccal root; DP,distal palatine root; M,mesial root.The solid line frame indicates the distal tension area, and the dashed line square indicates the mesial compression side. (H-L) H-E staining of the mesial side and the distal side of PDL between the untreated side and OTM side on day 7. n = 4 for each group (∗p < 0.05, ∗∗p < 0.01and ∗∗∗p < 0.001). Scale bars = 300 μm (D),500 μm (E),160 μm (G, J),40 μm (H, I, K, L). |
3.2. Mapping the distribution of Col1+ cells in the PDL under the orthodontic force
For further investigation, we conducted TRAP staining and immunohistochemical staining of paraffin sections of bilateral maxillary alveolar bone from mice after 7 days of OTM, a time point when substantial tissue structural alterations occur. TRAP staining was used to evaluate osteoclast expression by labeling with anti-tartrate acid phosphatase, a specific marker enzyme of osteoclasts. The results indicated significantly greater osteoclast accumulation around the mesial side of the PDL of the first molar in mice on day 7 than on the untreated side. However, no significant change was observed on the distal side (Fig. 2A). We used immunohistochemistry to label osteopontin (OPN), a secretory protein with crucial roles in bone mineralization and remodeling. Significant accumulation of OPN was observed around the distal side of the PDL on day 7, as compared with the untreated side, whereas no significant change was found on the mesial side (Fig. 2B). These results correspond to previous findings indicating that tooth movement is an interactive process in which osteoclastogenesis and osteogenesis occur together and are co-modified.
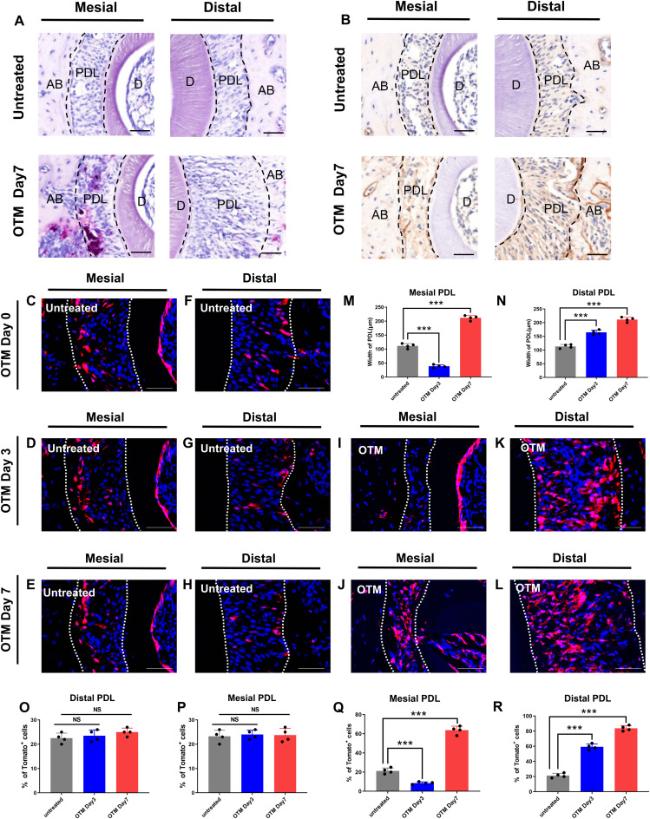
Fig. 2. Distribution of Col1+ cells in the PDL. (A) Tartrate-resistant acid phosphatase (TRAP) staining is peformed in PDL on the untreated side and OTM side on day7. (B) The expression of OPN is peformed in PDL on the untreated side and OTM side on day7. (C-L) Expression of Col1/Tomato+ cells at the mesial side and distal side of PDL during OTM. (M, N) Graphs show the width of PDL during OTM. (O-R) Graphs show percentiles of Col1+cells within the PDL of the mesial and distal side during OTM on Day 0,3,7. n = 4 for each group (NS p > 0.05, ∗∗p < 0.01and ∗∗∗p < 0.001). Scale bars = 40 μm (A,B), 50 μm (C-L). |
Next, we asked how Col1+ cells in the periodontal tissue might be altered under mechanical force. We sought to examine the distribution of Col1+ cells under the orthodontic force. To ensure the reliability and stability of Tomato expression in the inducible knockout mice, we compared the expression of Col1/Tomato+ cells on the untreated side at different time periods (0, 3, and 7 days) after OTM. We observed that the number of Col1+ cells in the PDL on the untreated side remained constant over time (Fig. 2C-H). Therefore, it is appropriate to select the tissue slices of WT mice injected with tamoxifen as the control group in the subsequent immunofluorescence experiment. Next, we traced the lineages of Col1+ cells in the PDL on both the untreated and OTM sides on day 0, 3, and 7. On the mesial side, after 3 days of OTM, narrowed width of the PDL as well as decreased number of Col1+ cells could be observed (Fig. 2I,Q). On the contrary, width of the distal PDL and the number of Col1+ cells increased (Fig. 2K,R). On day 7, width of the mesial PDL was found to expand since the third day but remained narrower than that on the untreated side (Fig. 2J,M). Similarly, although the number of Col1+ cells increased with respect to day 3, the number of cells remained far fewer than that on the untreated side (Fig. 2J,Q). These results suggested that the number of Col1+ cells increased proportionally with the application time of mechanical force. On the basis of these results, we concluded that Col1+ cells are a group of stem cells that respond to mechanical regulation.
3.3. Col1+ cells are associated with angiogenesis
Previous studies have demonstrated substantial spatial proximity between mesenchymal stem cells and blood vessels [25]. Additionally, the ability of mesenchymal stem cells to stimulate angiogenesis has been confirmed through numerous in vitro experiments and tissue engineering applications [26]. Consequently, we focused on determining whether the group of Col1+ cells that respond to mechanical force participated in angiogenesis. Endomucin(EMCN) is a classic endothelial cell marker that can be used to indicate the location of the blood vessels. EMCN was labeled on frozen sections of mouse alveolar bone through an immunofluorescence technique, thus revealing the presence of blood vessels in the PDL. Interestingly, on the untreated side of the PDL, only several Col1+ cells were observed, along with vascular tissue at both the mesial and distal side (Fig. 3A,D,G). On day 3, the number of blood vessels and Col1+ cells decreased in the mesial PDL and increased in the distal PDL. Additionally, a substantial number of Col1+ cells were detected adjacent to blood vessels on the distal side (Fig. 3B,E,H). On day 7, significantly more EMCN+ cells along with Col1+ cells could be observed in the distal PDL than the untreated side; the cell numbers were proportional to the duration of orthodontic force (Fig. 3C,F,I). In addition, the number of Col1+ cells on the mesial side of PDL on day 7 was greater than that on day 3 (Fig. 3B and C). These results indicated that both vascular endothelial cells and Col1+ cells proliferated under mechanical force, and the two cell types were in spatial proximity, thus suggesting that Col1+ cells may be closely associated with angiogenesis.
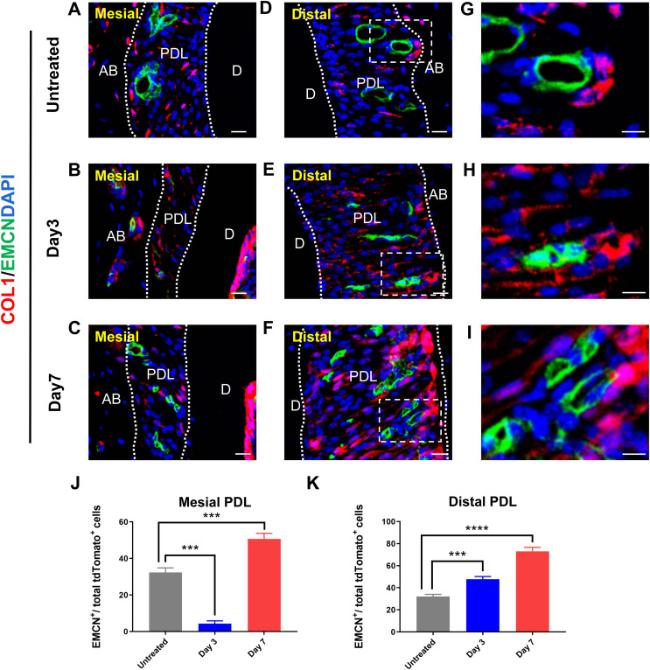
Fig. 3. Col1+ cells are associated with angiogenesis. (A-C) Immunofluorescence shows the distribution of Col1+ cells at the mesial side of PDL at various time points during OTM. (D-I) Immunofluorescence shows the distribution of Col1+ cells at the distal side of PDL at various time points during OTM. (J, K) Graphs show number of EMCN+ cells within each field view at various time points in the mesial and distal side of PDL after OTM. n = 3 for each group. (NS p > 0.05, ∗∗p < 0.01, ∗∗∗p < 0.001 and ∗∗∗∗p < 0.0001). AB, alveolar bone; D, dentin; PDL, periodontal ligament. Scale bar = 25 μm (A-F),10 μm (G-I). |
3.4. Col1+ cells can differentiate into fibroblasts under orthodontic force
Fibrous tissue is a major component often expressed throughout the PDL. We therefore used Periostin(POSTN) as a marker for fibroblasts, and conducted immunofluorescence staining to investigate the effects of mechanical regulation on fibrous tissue and the correlation between POSTN+ and Col1+ cells. Whereas POSTN+ cells were abundant in the PDL, fewer Col1/Tomato+ cells expressing POSTN were observed on the untreated side (Fig. 4A,D,G). On day 3, less double-positive cells expressing tdTomato and POSTN were observed among the total Col1/Tomato+ cells (Fig. 4B,E,H). After 7 days of OTM, significantly more double-positive cells were observed on both mesial and distal sides of the PDL than on the untreated side (Fig. 4C,F,I,J,K). These results indicated that Col1+ cells can differentiate into fibroblasts under mechanical force, thus supporting the structure and morphology of the periodontal tissue.
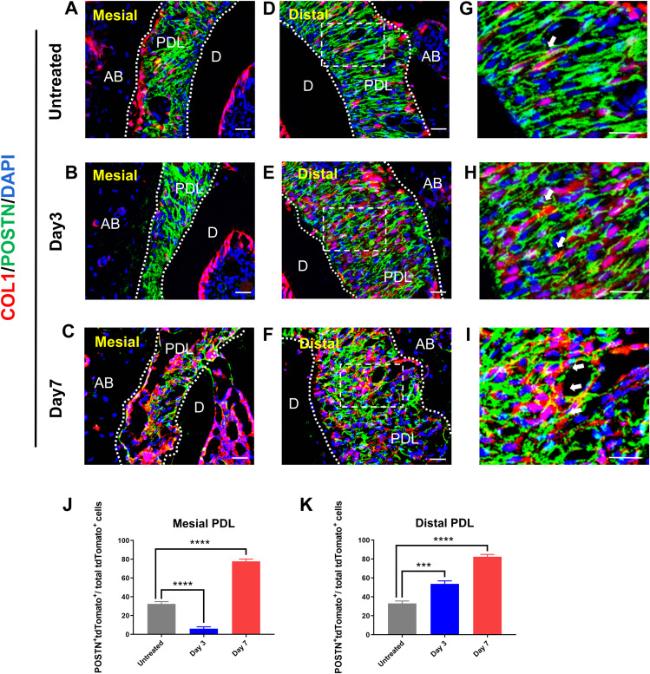
Fig. 4. Col1+ cells can differentiate into fibroblasts during OTM. (A-C) Immunofluorescence shows the distribution of Col1+ cells and POSTN+ cells at the mesial side of PDL at various time points during OTM. (D-I) Immunofluorescence shows the distribution of Col1+ cells and POSTN+ cells at the distal side of PDL at various time points during OTM. Arrows indicated POSTN+ and Col1+ cells. (J, K) Graphs show number of Col1+ cells as well as POSTN+ cells within each field view at various time points in the mesial and distal side of PDL after OTM. n = 3 for each group. (NS p > 0.05, ∗∗p < 0.01, ∗∗∗p < 0.001 and ∗∗∗∗p < 0.0001). AB, alveolar bone; D, dentin; PDL, periodontal ligament. Scale bar = 25 μm (A-I). |
3.5. Col1+ cells contribute to the osteolineage under orthodontic force
The production of a large quantity of new bone matrix is a notable characteristic of bone tissue remodeling under mechanical regulation. As a classic zinc-fingered osteoblast-specific transcription factor, osterix (OSX) is essential for osteoblast proliferation, differentiation, and bone formation. Through OSX labeling of mouse alveolar bone and the use of immunofluorescence techniques, we identified the relationship between Col1+ cells and osteogenesis. On the untreated side, OSX was moderately expressed in the PDL (Fig. 5A,D,C). On day 3, OSX+ cells in the periodontal membrane increased in the distal PDL while decreased in the mesial PDL (Fig. 5B,E,H). On day 7, OSX+ cells increased significantly in both mesial and distal side of PDL (Fig. 5C,F,I). OSX is an exocrine protein expressed primarily in cell membrane and extracellular matrix. The colocalization of Col1+ cells with OSX suggested that Col1+ cells had the potential to differentiate into osteoblasts in a time-dependent manner (Fig. 5D-I).
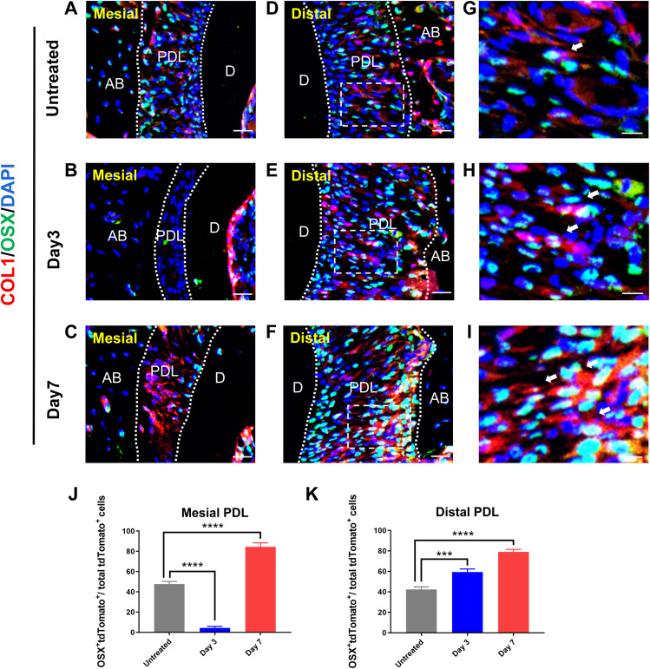
Fig. 5. Col1+ cells contribute to the osteolineage during OTM. (A-C) Immunofluorescence shows the distribution of Col1+ cells and OSX+ cells at the mesial side of PDL at various time points during OTM. (D-I) Immunofluorescence shows the distribution of Col1+ cells and OSX+ cells at the distal side of PDL at various time points during OTM. Arrows indicated OSX+ and Col1+ cells. (J, K) Graphs show number of Col1+ cells as well as OSX+ cells within each field view at various time points in the mesial and distal side of PDL after OTM. n = 3 for each group. (NS p > 0.05, ∗∗p < 0.01, ∗∗∗p < 0.001 and ∗∗∗∗p < 0.0001). AB, alveolar bone; D, dentin; PDL, periodontal ligament. Scale bar = 25 μm (A-F),10 μm (G-I). |
4. Discussion
Advances in medicine and increased health consciousness among individuals have led to a growing demand for the diagnosis and treatment of dental and maxillofacial abnormalities. Consequently, orthodontists have closely examined methods to safely and efficiently shorten the duration of orthodontic treatment while minimizing the likelihood of recurrence and complications. The fundamental principle of OTM is the reconstruction of periodontal tissues, in which MSCs play a crucial role [27]. Previous research has demonstrated that MSCs detect mechanical forces and can differentiate into various cell lineages [28,29]. However, the microenvironment of MSCs in vivo is complex, and in vitro studies do not fully reflect their biological behavior [30,31,32]. In order to study the role of MSCs in regulating periodontal remodeling under orthodontic force, it is more appropriate to label and track MSCs in vivo.
This technology has also been a powerful tool allowing our research group to advance understanding of the mechanism of tooth movement. However, limited reports have described the use of lineage tracing technology in OTM.
Lineage tracing is a technique used to mark and identify a single cell and all its progeny cells, and consequently explore cell lineage. This method enables tracking and observation of the number, location, and differentiation of progeny cells [33]. The Cre-loxP system, a widely used recombinant enzyme system in lineage tracing technology [14], involves insertion of the LOXP-STOP-LOXP sequence in front of a reporter gene. When the Cre recombinase is activated, it cuts the STOP sequence in the inserted sequence, thus resulting in reporter gene expression in all cells expressing Cre recombinase, and their progeny cells. Additionally, Cre recombinase binds mutated ER (CRE-ER) [34]. When tamoxifen is administered, CRE-ER enters the nucleus and initiates recombination, thus resulting in the labeling of an inducible specific fluorescent protein. In recent years, induced cell lineage tracing technology has become an essential tool for studying the generation and outcome of stem cell lineages [35].
In previous studies, some researchers have used axis inhibition protein 2 (Axin2), Gli1, and Prrx1 to mark mesenchymal stem cells [36,37,38]. Gli1+ and Prrx1+ cells play important roles in periodontal membrane remodeling and molar growth in mice. The dynamic distribution and differentiation tendency of Col1+ cells under mechanical force have not been reported. Our study show that Col1+ cells, which are sensitive to mechanical force, are mesenchymal stem cells that distributed in periodontal tissues such as dental pulp, periodontal membrane, and alveolar bone. The OTM mouse model is commonly used for examining periodontal tissue remodeling through mechanical regulation. We show that during OTM, the number of Col1+ cells and their proliferative ability increased. Furthermore, through immunofluorescence and other techniques, we show that Col1+ cells have the potential to stimulate angiogenesis, osteogenesis, and fibroblast differentiation under mechanical force.
In this study, we successfully created a mouse tooth movement model by using the induced Col1+ cell lineage tracer. We confirmed that the Col1+ cell lineage participates in the periodontal remodeling of OTM, and we detected markers of osteogenesis, angiogenesis, and fibroblasts by immunofluorescence technology. EMCN is a mucin-like sialoglycoprotein that can indicate the location of blood vessels in the periodontal membrane by labeling vascular endothelial cells [39]. POSTN is an osteoblast-specific protein factor with relatively high levels in collagen-rich connective tissues such as heart valves, tendons, and periodontal ligaments [40]. Osterix, a novel zinc-containing finger transcription factor, has long been recognized as an essential transcription factor for osteoblast differentiation and bone mineralization [41]. Here, we used immunofluorescence technique to explore the osteogenic and fibrogenesis potential of Col1+ cells and its relationship with angiogenesis by labeling EMCN+, POSTN+, and OSX+ cells. Our findings demonstrated that Col1+ cells have the potential to stimulate angiogenesis, osteogenesis, and fibroblast differentiation under mechanical force. Importantly, MSCs are widely distributed, are heterogeneous, and exist in different functional subgroups. In the future, single-cell sequencing technology will be used to group MSCs through transcriptomics, thus enabling identification of tissue-specific markers and providing new insights into periodontal tissue regeneration. Furthermore, the Col1-CreERT2; R26tdTomato mice constructed in this study could be used to track important biological processes and lineage outcomes involved in Col1+ cells, thus facilitating analysis of periodontal injury and periodontitis models.
In conclusion, this study successfully used lineage tracer technology combined with the OTM model to achieve spatiotemporal specific labeling and tracking of Col1+ cell differentiation under orthodontic force. This work may provide a novel animal model and research strategy for studying the mechanism of tooth movement.
Author contributions
Xinyu Wang, contributed to conception and design, data acquisition and interpretation, drafted and critically revised the manuscript;
Xiangru Huang, contributed to analysis and interpretation, drafted and critically revised the manuscript;
Xin Gao, contributed to analysis and data interpretation, critically revised the manuscript;
Hongyuan Xu, Anting Jin, Xijun Wang, contributed to interpretation, critically revised the manuscript;, Siyuan Sun,Yuanqi Liu, Yanfei Zhu, Jingyi Liu, Tingwei Lu, contributed to acquisition, critically revised the manuscript; Qinggang Dai, contributed to conception, and data acquisition, critically revised the manuscript; Lingyong Jiang, contributed to conception and data design, acquisition, drafted and critically revised the manuscript. All authors gave their final approval and agree to be accountable for all aspects of the work.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported in part by grants from the National Natural Science Foundation of China (82071083, 82271006, 82101048); the National Science Foundation of Shanghai (21ZR1436900, 22ZR1436700); the Program of Shanghai Academic/Technology Research Leader (20XD1422300); the Cross-Disciplinary Research Fund of Shanghai Ninth People's Hospital, Shanghai Jiao Tong University School of Medicine (JYJC202116, JYJC201902); the Clinical Research Plan of SHDC(SHDC2020CR4084); the Biomaterials and Regenerative Medicine Institute Cooperative Research Project Shanghai Jiao Tong University School of Medicine (2022LHB02); the Project of Biobank of Shanghai Ninth People's Hospital, Shanghai Jiao Tong University School of Medicine (YBKB201909, YBKB202216); the Research Discipline Fund no. KQYJXK2020 from Ninth People's Hospital, Shanghai Jiao Tong University School of Medicine, and College of Stomatology, Shanghai Jiao Tong University; the Original Exploration Project of Shanghai Ninth People's Hospital, Shanghai Jiao Tong University School of Medicine (JYYC003); the Two-Hundred Talent Project of Shanghai Jiao Tong University School (20221809); Shanghai Science and Technology Innovation Action Plan-International Science and Technology Cooperation Program (23410713600).
Ethics statement
This study complied with all relevant ethical regulations for animal testing and research. All experimental animal procedures were approved by the Institutional Animal Care and Research Advisory Committee of the Shanghai Ninth People's Hospital, School of Medicine, Shanghai Jiao Tong University and were performed according to the institutional guidelines (approval number: SH9H-2023-A92-1) and the ARRIVE (Animal Research: Reporting In Vivo Experiments) guidelines. All mice were bred and maintained under specific pathogen-free (SPF) conditions.
Declaration of competing interest
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Acknowledgment
The authors thank the Laboratory for Digitized Stomatology and Research Center for Craniofacial Anomalies of Shanghai Ninth People's Hospital for assistance.The authors also thank the Shanghai Institute of Precision Medicine for bioimaging and molecular experiment support.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}