
1. Introduction
Glioblastoma (GBM) is the most aggressive type of brain tumor that originates from glial cells and is characterized by a high fraction of cancer cells with stem-like properties in their tumor microenvironment [1]. This subpopulation of cancer cells, referred to as “cancer stem cells” (CSCs), are defined by their self-renewal and tumor-initiating capacities, and are tightly associated with tumorigenesis, metastasis, high resistance to cancer treatments, and poor survival rate in patients [2,3]. CSCs are often characterized based on expression of several putative stem cell surface markers (e.g., CD133), cytoplasmic proteins (Nestin), and nucleus proteins (Nanog, Oct4) [4,5,6,7]. For example, expression of CD133 and Nestin have been found to increase with increasing tumor grade in glioma compared to normal brain tissues, and are associated with worse overall survival in glioma patients [8]. Suppression of Nanog in GBM was shown to ameliorate drug resistance by inducing cell cycle arrest of CSCs [9].
Despite their prognosis significance, the origin of CSCs in GBM is yet fully understood. The emergence of CSCs could originate from both intrinsic and extrinsic factors. Genetic variation, epigenetic alternation, and the epithelial-to-mesenchymal transition (EMT) in tumor cells are the most common cell-intrinsic factors found to drive emergence of CSCs, while cell-extrinsic factors usually arise from aberrant biochemical and mechanical cues in the tumor microenvironment [10,11]. Beside changes in biochemical cues such as oxygen, pH, and immunosuppressive properties [12,13,14], the role of tissue mechanics in the aggression, invasion, and stemness of cancer cells have also been extensively appreciated. The transformation and properties of CSCs are tightly regulated by the biophysical properties of their surrounding microenvironment, including but not limited to mechanical confinement, geometrical topography, solid stress, shear stress, and matrix stiffness [15,16,17,18,19,20,21,22]. Influenced by their extracellular microenvironment, CSCs have shown distinct mechanobiological properties, such as cell adhesion [23], deformability [24,25,26], and contractility [27] from non-CSCs and vary depending on the type of cancer. For example, GBM CSCs tend to experience higher matrix stiffness and myosin-dependent contractility [28], while CSCs in inflammatory breast cancer resembling an epithelial-like phenotype are less stiff and have lower cellular contractility [23].
A major mechanical characteristic of GBM is the accumulated stress inside the tumor due the physical confinement of the tumor by the rigid skull. In this scenario, GBM cells in the overcrowding tumor microenvironment undergo mechanical constraints that impact both the cell body and nucleus, leading to numerous cellular alterations such as cell deformability and traction force as well as cancer cell DNA transcription [20,29,30]. In addition, gradients of mechanical stresses generated within the confined tumor microenvironment can mechanically induce a malignant phenotype in cancer cells at different regions, influence the emergence and spatial distribution of CSCs within the tumor. Recent studies have found that distribution of CSCs in tumor tissue is not uniform both in vitro and in vivo. For instance, the EMT and CSCs preferentially occur in the peripheral regions of cell monolayers [27,31,32]. Although plenty of evidence support that these mechanical cues in the tumor microenvironment influence CSC phenotype and mechanics, it remains unclear how cooperative intracellular and intercellular forces due to mechanical constraints in tumor regulate the emergence and spatial patterning of CSCs in GBM. More importantly, little is known about the underlying mechanosensitive signaling that initiates CSC emergence and maintains the spatial heterogeneity of CSC in GBM tumor.
Mechanical signals within the tumor microenvironment are transduced into tumor cells by multiple receptors, such as integrins, G-protein-coupled receptors, and mechanosensitive ion channels [33]. Several studies have confirmed integrin-mediated activation of the Rho-family GTPases [34], FAK/PI3K/AKT [35] and ERK/MAPK pathways [36] - each of which is enhanced in cancer cells in response to mechanical cues. Study has found that interfacial geometry initiated the differentiation of cancer cells into CSCs through integrin α5β1, MAPK and STAT pathways, thus promotes tumorigenicity in melanoma and human prostate cancer [31]. Moreover, receptors anchored to the actin cytoskeleton, including integrins and focal adhesion can rapidly relay mechanical stimuli through release of actin-bound transcription factors and direct coupling to the nuclear membrane thus mediate CSC phenotype in GBM tumor [37,38]. Among the mechanosensitive ion channels, Piezo1 tension-gated channels have recently become a topic of interest and have been implicated in the regulation of CSCs. Studies have shown that Piezo1 expression is elevated in CSCs compared to non-CSCs [39,40], and that mechanosensing by Piezo1 regulates the stemness and self-renewal of CSCs through modulation of various signaling pathways involved in cell survival, proliferation, and differentiation [41]. Confinement sensing via Piezo1/PKA and Myosin II pathways that optimize cell motility has been discussed in melanoma model [42]. Recent study has also revealed a feedforward mechanism mediated by Piezo1 ion channels and tissue mechanics that promote glioma aggression [43]. Yet, understanding of the mechanism by which Piezo1 “senses” mechanical constraints experienced by GBM cells in the tumor microenvironment and how Piezo1 mechanosensitive signaling influences the phenotypic switch of cancer cells into CSCs still require further exploration.
In this study, we use a two-dimensional (2D) micropatterned multicellular model to examine the impact of mechanical constraints in tumor on the emergence and spatial patterning of CSCs in GBM. We specifically investigated how the interplay between forces arisen from the cell-ECM and cell-cell interactions guide the emergence and spatial distribution of CSCs in GBM. In different geometric multicellular patterns, GBM cells in the peripheral regions are all found to express a higher level of CSC makers. The distinct spatial pattern of CSCs was found to correspond to the gradients of mechanical stresses generated within the confined environment. Our study further highlighted the coordination among Piezo1 mechanosensitive channels, integrins, and cadherins in regulating GBM cell mechanosensing, and demonstrated that the upregulation of Piezo1 plays a critical role in the phenotypic switch of GBM cells to CSCs. These findings highlight the role of mechanical forces that originate from cellular contractility and intercellular interaction, emerged from the mechanical constraints in the multicellular organization, in determining the unique spatial patterns of CSCs in GBM.
2. Materials and methods
2.1. Cell culture and reagents
Mouse glioma cell line CT-2A (Milipore Sigma) with characteristics of GBM phenotypes were used. CT-2A cells were cultured in Dulbecco's modified Eagle's medium (DMEM, Sigma-Aldrich) supplemented with 10% heat-inactivated fetal bovine serum (FBS, Gibco), 2 mM glutamine (Gibco) and 1% penicillin/streptomycin (Gibco) in a 37 °C and 5 % CO2 incubator. Fresh media was replaced every 2 days, and all cells were cultured for no more than 6 passages.
2.2. Fabrication of polyacrylamide (PA) hydrogel substrates
PA hydrogels were prepared according to a previously reported protocol [44]. Prior to gel being cast, glass coverslips (22 mm, Fisher Scientific) were attached to a Petri dish with a punctual hole of 13 mm diameter using Polydimethylsiloxane (PDMS, Dow Corning) of mass ratio 10:1 (Base Elastomer:curing agent) and baked in an 80°C oven. The coverslips attaching to petri dishes were then treated with 0.1 M sodium hydroxide (NaOH, Sigma-Aldrich) solution and were heated till the entire coverslips were covered with a uniform monolayer of NaOH. NaOH-coated coverslips were further functionalized with 3-aminopropyltriethoxysilane (Sigma-Aldrich) for 5 min and then rinsed three times with distilled water. The coverslips were then incubated with 0.5% glutaraldehyde (Sigma-Aldrich) for 30 min, washed three times with distilled water, and then dried at room temperature. An 18 μl mixture of 40% acrylamide, 2% bis-acrylamide (Sigma-Aldrich), 0.1% ammonium persulfate (APS, Sigma-Aldrich), and 0.1% tetramethylethylenediamine (TEMED, Sigma-Aldrich) were added to the center of the coverslips, and then covered by another 18 mm coverslip that was hydrophobically treated with Dichlorodimethylsilane (Sigma-Aldrich). To prepare PA gels for traction force microscopy, 0.2-μm, yellow-green fluorescent (505/515) microbeads (Thermo Fisher Scientific) were added to the 18 μl gel solution and mixed thoroughly before adding to the coverslips. After 60 min of polymerization, the top 18 mm coverslips were peeled off to have the PA gel attached on the 22 mm coverslips attaching to petri dishes.
2.3. Microcontact printing on PA hydrogel substrates
PA gel substrates were first activated by treating with 200 μL of 1 mM Sulfo-SANPAH (sulfosuccinimidyl 6-(4′-azido-2′-nitrophenylamino)hexanoate, Pierce) in 50 mM HEPES (N-2-hydroxyethylpiperazine-N-2-ethane sulfonic acid, Gibco) buffer under ultraviolet (UV) light for 20 min followed by washing with phosphate-buffered saline (PBS) thrice, and were then dried at room temperature. PDMS stamps with different geometric patterns were first fabricated using standing photolithography and replica molding [45]. PDMS stamps were functionalized with a fibronectin solution (50 μg/ml; Sigma-Aldrich) for 60 min at room temperature, then were put into conformal contact with activated PA gels for 1 min and peeled off. PA gels functionalized with different patterns of fibronectin proteins were then washed with PBS twice and stored in PBS for experiments within one week. Prior to cell loading, functionalized PA gel substrates were sterilized under UV light for 15 min in PBS and for 15 min in cell culture media. 15,000 CT-2A cells were then seeded onto each sterilized PA gel substrate. These loaded cells would attach to the micropatterned adhesive areas and be cultured for 3 days in an incubator to form different geometric patterns of cell monolayers.
2.4. Traction force microscopy
To measure traction forces generated from cells in the 2D cell monolayer, fluorescent microbeads embedded in PA gels underneath the patterned cells were imaged firstly to recode the position of each microbead using a Zeiss Observer microscope with a 40 × objective. Then cells cultured on PA gels were lysed by adding 100 μL of 1 N NaOH to the cell culture dish, and fluorescent images of the same locations were taken again to record the positions of these fluorescent microbeads as a control. The two sets of fluorescence microscopic images prior and post cell lysis and a phase contrast image of cells were used to measure bead displacement using particle image velocimetry. The resultant displacements were then used to calculate the traction force using Fast Fourier transform cytometry [46].
2.5. Immunofluorescence staining and microscopy
Cells were fixed with 4% Paraformaldehyde (Electron Microscopy Sciences) for 20 min and then permeabilized with 0.3% Triton X-100 (Roche Applied Science) for 10 min at room temperature. After permeabilization, cells were blocked with blocking buffer for 60 mins at room temperature. Primary antibodies (Table S1) were incubated with cells overnight at 4°C or 60 min at room temperature. Goat-anti mouse Alexa Fluor 488 and/or goat-anti rabbit Alexa Fluor 546 IgG secondary antibodies (Invitrogen) were used as secondary antibodies. The immunostained cells were imaged under an inverted fluorescent microscope (Zeiss Axio Observer.Z1).
3. Results
3.1. Mechanical constraints in tumor guide the spatial patterns of GBM CSCs
To mimic the mechanical constraints experienced by cancer cells, we employed a 2D multicellular model of GBM by micropatterning of mouse GBM cells (CT-2A, Sigma Aldrich) into geometrically confined monolayers (Fig. 1a). Using a standard microcontact printing technique (Fig. S1) [45], mouse GBM cells were patterned into different shaped monolayers on PA hydrogel substrates with an elastic modulus of 3 kPa (Fig. 1b). PA gels were embedded with fluorescent microbeads to measure the traction force distribution within micropatterned multicellular monolayers [47]. We first examined the spatial distributions of CSCs in 500 μm-diameter circular GBM cell colonies by measuring expressions of key CSC makers [48], including canonical self-renewal transcription factors (Nanog, Oct4), CSC membrane marker (CD133) and intermediate filament marker (Nestin) after 3 days of cell culture (Fig. 1c). The fluorescent intensity of CSC makers on each single cell in a 2D colony after immunostaining was measured and presented as a heat map (Fig. 1d). The CSC surface maker analyses showed non-uniform spatial distributions of GBM CSCs in the micropatterned colonies, whereas GBM cells that reside in peripheral regions showed elevated expressions of all four CSC markers (Nanog, Nestin, Oct4, and CD133) compared to the those in the proximal regions (Fig. 1e-f).
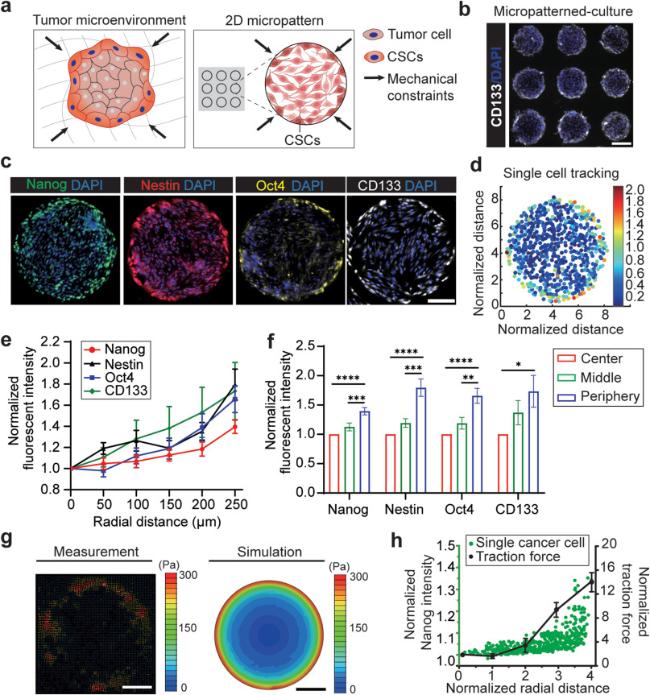
Fig. 1. Mechanical constraints in tumor guide the spatial patterns of GBM CSCs. |
To gain a deeper understanding of the mechanical factors that drive the transition of GBM cells into CSCs, we employed fluorescent bead-based traction force microscopy [47] to map the cellular traction force distributions within the micropatterned multicellular monolayers (Fig. 1g). The force analyses showed a strong correlation between the spatial distributions of CSCs and traction force in the micropatterned GBM colonies. In consistent with the measured trends of CSC marker expressions, traction forces generated by GBM cells increased from the central regions to the peripheral regions of the circular patterns (Fig. 1h). Such gradients of cellular force distributions in the patterned cell colonies can be explained by the unique mechanical constraints due to geometric confinement, as predicted by a finite-element model of multicellular sheet mechanics (Fig. 1g) [49]. We further investigated whether varying geometries of the multicellular patterns would alter the spatial distributions of CSCs and force maps. The same phenomenon was observed in all types of sizes (100, 300, and 500 μm-diameter circular) and geometries (circular, rectangular, T-shape) of cell patterns, whereas CSCs emerged more in the peripheral regions than in the central regions of micropatterned GBM cell monolayers (Fig. S2a-b). Moreover, traction forces generated by cells in the peripheral regions were all significantly higher than those in the inner regions (Fig. S2c-d). Meanwhile, such spatial CSC distribution was not observed under a nonconfined condition, indicating that geometric confinement is an inductive cue for emergence of CSCs (Fig. S2a-b).
3.2. Cell-cell and cell-ECM interactions mediate the force-driven spatial patterns of CSCs
We explored how the interplay between the cell-cell interaction and cell-ECM interaction under mechanical constraints mediates the force-driven emergence and spatial patterning of CSCs in the geometric confined GBM cell colonies. As cells in the GBM colonies interact with surrounding cells and ECM, mechanical signals are transmitted by cell adhesive molecules including integrins and cadherins to actomyosin cytoskeleton, thus regulate cellular forces and downstream mechano-responsive signaling [33] and mediate CSC phenotype in GBM [37,38]. We conducted immunostaining to assess the expression of cell adhesion molecules that are crucial for cell-cell adhesion (E-cadherin, N-cadherin, P-cadherin) and cell-ECM adhesion (integrin α5β1) (Fig. 2a). The results showed that the expressions of N-cadherin and P-cadherin increased significantly from the center to the peripheral regions of the patterned cell colonies, while E-cadherin expressed uniformly in the colonies (Fig. 2b). Such findings were expected, since E-cadherin expression has been found to be scarce in glioma cells, while N-cadherin upregulation has been widely exploited in glioma prognosis [50]. In addition, integrin adhesome takes a critical role in transmitting mechanical signals from the ECM to the cytoskeleton and regulating actomyosin-mediated traction force exert by the cells on the ECM components [51]. Previous studies have revealed that abnormal expression of integrin α5β1 in GBM cells results in a pathological alteration of cell-ECM interactions, and impact angiogenesis, migration, proliferation and therapy resistance of GBM [52,53,54]. Our results proved that integrin α5β1 expression in the patterned cell colonies increased significantly from the center to the peripheral regions, which implicated the corresponding force gradients in the patterned GBM colonies (Fig. 2a-b). The regression analysis between expression of adhesion molecules (E-cadherin, N-cadherin, P-cadherin, and integrin α5β1) and the CSC marker Nestin further showed that the expression level of Nestin in GBM cells in the colony is positively correlated with the expression levels of these adhesion molecules N-cadherin, P-cadherin, and integrin α5β1 but not E-cadherin (Fig. 2c).
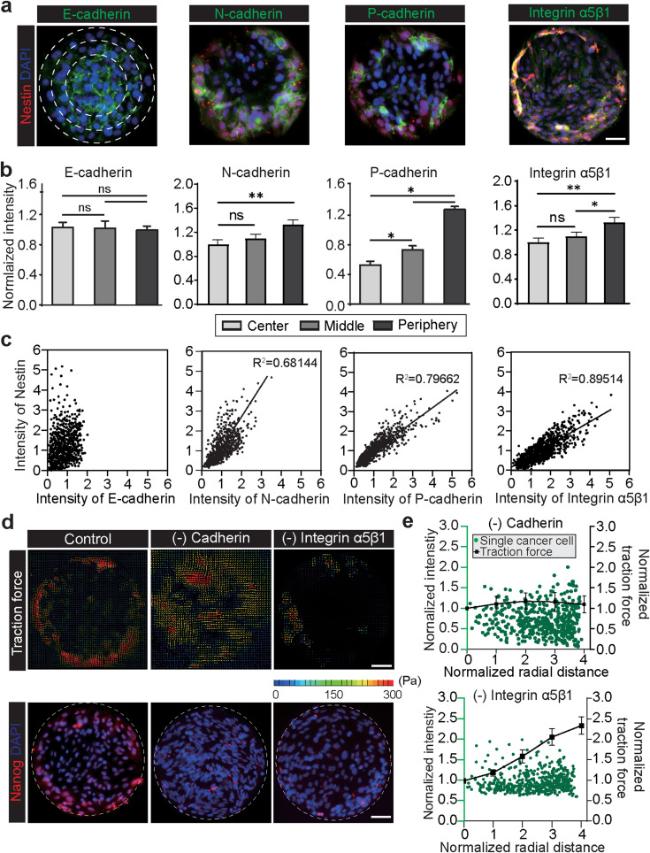
Fig. 2. Cell-cell and cell-ECM interactions mediate the force-driven spatial patterns of CSCs. |
Furthermore, we utilized pharmacological treatments to interrupt the cell-cell and cell-ECM interactions to delve into the mechanotransductive mechanisms underlying the emergence and spatial patterning of CSCs in the geometric confined GBM colonies. We used a low calcium medium, which effectively suppressed the adhesion activities of N-cadherins and P-cadherins, thereby interrupting the cell-cell interactions [55]. The traction force microscopy measurements revealed that disrupting cadherins greatly attenuated the traction force gradient in the GBM colonies, while the total traction force remained the same (Fig. 2d-e). We also applied an integrin α5β1 antibodies to block the cell-ECM adhesion and to disrupt the traction force that the cells exert on the PA substrate [31]. The force analyses suggested that while inhibition of integrin α5β1 reduced the overall magnitude of traction forces in most cells, the force gradient remains in the GBM colonies (Fig. 2d-e). Moreover, we directly inhibited cellular traction forces with Blebbistatin (20 μM), an inhibitor of myosin-II-specific ATPase [54]. Inhibition of actomyosin force also attenuated the force gradient in the GBM colonies (Fig. S3a-c). We then checked whether disrupting the mechanical constraints in the geometric confined GBM colonies would attenuate the emergence and spatial distribution of CSCs. Blocking cell-cell, cell-ECM interactions and traction force reduced the overall percentages and attenuated spatial distribution of CSCs in the GBM colonies of according to all four CSC markers (Nanog, Nestin, Oct4, and CD133) (Fig. 2e, Fig. S3d). Overall, our results suggest that the emergence and spatial patterning of CSCs require the development of gradient distribution of traction force mediated both by the cadherin-mediated cell-cell interaction and integrin-mediated cell-ECM interaction.
3.3. Force-driven emergence of GBM CSCs is mediated by mechanosensitive Piezo1 ion channels
We next explored the Piezo1-mediated mechanotransductive mechanisms underlying the force-driven emergence and spatial patterning of CSCs in the geometric confined GBM cell colonies. Upregulation of Piezo1 mechanosensitive ion channel has been implicated in the growth of GBM stem cells and tumor development [41]. A previous study has also revealed a feedforward mechanism mediated by Piezo1 and integrin-focal adhesion kinase (FAK) signaling that promote glioma aggression by remodeling tumor mechanics [43]. We first examined the expression of Piezo1 in the 2D confined GBM cell colonies (Fig. 3a) and found that Piezo1 was prominently concentrated in the peripheral regions of the micropatterned colonies (Fig. 3b). As indicated in Fig. 3c, there is a positive correlation between Piezo1 expression and CSC marker Nanog, a transcription factor that promote GBM malignant phenotype [56]. Upon its activation by either mechanical or chemical stimulator, Piezo1 opens and allow influx of calcium ion (Ca2+) into the intracellular domain and regulate of cellular contraction [57]. We transfected the cell monolayer with Fluo4 AM, a calcium indicator, and found that calcium mostly concentrated in the peripheral regions of the micropatterned colonies (Fig. 3d-e). The transient activities of calcium upon Piezo1 activation were also tested using Yoda1, a Piezo1 chemical agonist [58]. Upon addition of Yoda1, we observed an immediate rise in intracellular calcium in all regions of the GBM colonies (Fig. 3f). However, the rise in intracellular calcium in cells in the peripheral regions was still significantly higher compared to cells in the central regions, which could be explained by the overexpression of Piezo1 in the peripheral regions. From these results, we confirmed that Piezo1 signaling plays a critical role in regulating GBM CSC phenotype.
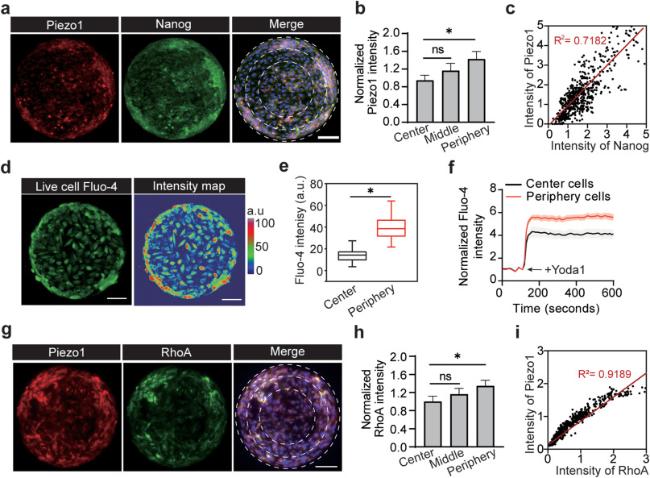
Fig. 3. Force-driven emergence of GBM CSCs is mediated by mechanosensitive Piezo1 ion channels. |
Next, we tested whether focal adhesions and RhoA activities collaborate with Piezo1 to drive traction force elevation in GBM cells and CSC phenotype switch. This spatially segregated Piezo1 channel activity promotes efficient assembly and signaling of integrin-mediated focal adhesions, which serve as highly localized subcellular domains for Piezo1 to perceive mechanical stimuli at both the cell surface and the cell intracellular structures [43,59]. As an integrin-interacting molecule, Piezo1 also took part in assembly of focal adhesion [59]. From the co-staining of Piezo1 and FAK, we observed that Piezo1 colocalizes with FAK in individual CSCs (Fig. S4). Moreover, we found a strong correlation between Piezo1 expression and RhoA activities in the GBM cell colonies (Fig. 3g-i). RhoA is a downstream effector of intracellular calcium influx and initiate actin polymerization and thus generate actomyosin-mediated traction force [59]. Activation of Piezo1 and downstream RhoA activities mostly occur in the peripheral regions of the GBM cell colonies (Fig. 3i). These findings suggest that mechanosensation of the mechanically constrained microenvironment by Piezo1 triggers calcium influx in GBM cells, which in turn promotes RhoA activities to drive assembly of integrin-mediated focal adhesion and enhance traction force more in the peripheral regions of the cell colonies.
We next studied the interplay between integrin, cadherins, and Piezo1 in regulating mechanosensation of GBM cells to the mechanically constrained microenvironment (Fig. 4a). Interestingly, we first found that blocking of integrin α5β1 with antibodies significantly affected the overall Piezo1 expression and distribution in the micropatterned GBM cell colonies, blocking cadherins by using low calcium media did not alter Piezo1 expression in cells in the peripheral regions (Fig. 4b). To confirm whether Piezo1 recruitment precedes the change in traction force, we antagonized expression of Piezo1 using pharmacological treatment with 2.5 μM of GsMTx4 peptide for 1 h. Gsmtx-4 at μM concentration has been proven to inhibit Piezo1 mechanosensitive functions by 80% [59]. The efficacy of GsMTx4 was confirmed by a significant decrease in Piezo1 expression compared to the non-treated group, as demonstrated in Fig. 4c and d. We observed not only the overall intensity of Piezo1 decreased, but also the disappearance of the localization of Piezo1 at the edge of the circular GBM cell colonies. Furthermore, upon inhibition of Piezo1, we found the disruption in the distribution gradient of integrin α5β1, where the fluorescent intensity of integrin α5β1 in cells in the peripheral regions significantly decreased (Fig. 4d), implying the mutual interaction of Piezo1 and integrin adhesion during CSCs’ mechanosensing to mechanical constraints. Meanwhile, blocking Piezo1 did not affect N-cadherin activities among the GBM micropatterns and vice versa, as the intensity of N-cadherin in the cells located at the edge was still higher compared to those at the center region. Inhibition of Piezo1, along with the associated alleviation of integrin activities, resulted in the decrease in traction forces in GBM cell colonies and disappearance of the spatial patterning of CSC marker Nanog, confirming a Piezo1 force-driven mechanism in regulating the organization of GBM CSCs within the tumor (Fig. 4e-f).

Fig. 4. Antagonizing Piezo1 prevents emergence of GBM CSCs. |
4. Discussion
GBM tumor in vivo are mechanically constrained due to the geometric confinement of the skulls, and such mechanical constraints cause changes in various cell-ECM and cell-cell interactions that regulate the phenotype and functional behaviors of the cancer cell population. Our study specifically examined the impact of mechanical constraints in GBM tumor on the emergence and spatial patterning of CSCs in a 2D micropatterned multicellular model. Our results revealed that GBM cells in the peripheral regions of different geometric multicellular patterns experiencing more mechanical constraints tend to emerge into CSCs with elevated expressions of CSC markers (e.g., Nanog, Nestin, Oct4, CD133). Using hydrogel-based traction force microscopy, we found that the distinct spatial patterns of CSCs were positively correlated with the traction force gradients in the mechanically confined cell colonies. Our measurements indicated that the traction forces generated by cells in the peripheral regions are larger than those in the central regions of cell colonies, which is consistent with previous studies showing the dependency of traction force magnitude on localization of the cells in geometric patterns [27,31]. The 2D multicellular model although might not be perfect, is an effective strategy that can overcome the limitations of current 3D models and in vivo systems in assessing mechanical cues such as force gradient generated by the tumor cells, thus allow studying the mechanical constraints-mediated biophysical mechanism and signaling pathways of in GBM tumor. To extend the findings of this study, further experiments could be conducted in 3D models (such as cancer spheroids and organoids) and in vivo tumor samples integrated with new mechanosensing methods such as FRET-based molecular sensors [60,61] to validate how mechanical constraints regulate mechanical signals and spatial patterning of CSCs in GBM tumor.
Our study further explored the underlying mechanotransductive machinery involved in the mechanical constraints-mediated emergence and spatial patterning of CSCs in GBM. The gradients of traction force and spatial distribution of CSCs in the micropatterned GBM cell colonies were largely mediated by the collaborative mechanotransducive interactions between the integrin-mediated cell-ECM adhesions and cadherins-mediated cell-cell adhesions. In the presence of cadherin-based cell-cell interaction, the cell-ECM traction force increases at the peripheral of the confined cell colonies. Without cadherin signaling (weak cell-cell interaction), while the overall cell-EMC traction force remains the same, the force transmission among the cells in the confined colonies is disrupted, resulting in dissipated gradient of traction force distribution in the micropatterns. Meanwhile, blocking the actomyosin traction force and integrin-mediated cell-ECM adhesion all can attenuate the overall traction force magnitude that is necessary to provoke the force-dependent emergence of CSCs. This indicates that mechanical constraints in the geometrically confined environment influence actomyosin traction force, cell-cell adhesion, and cell-ECM adhesion relayed mechanotransduction, and thus regulate the spatial heterogeneity of cancer cells’ fate.
Our study highlighted a key role of Piezo1 mechanosensitive channels as the in regulating the mechanosensing and emergence of GBM CSCs in response to the mechanical constraints in the geometrically confined tumor environment. Mechanosensitive ion channels Piezo1 have been suggested as potential prognostic marker in gliomas for their ability to influence tumor cell tumorigenesis [62]. Our results showed heterogeneous expressions of Piezo1 in cells in different regions of micropatterned GBM colonies, which correspond to the gradients of traction force and spatial distribution of CSCs. We applied pharmacological treatment of GsMTx4 to Inhibit Piezo1 activities in the GBM colonies and found that attenuating Piezo1-medaited mechanosensing to these mechanical constraints signals successfully disrupted both the force gradient and spatial distribution of CSCs in the geometrically confined tumor environment. Our study further revealed interplay among Piezo1 mechanosensitive channels, integrins, focal adhesion, and downstream cytoskeletal machinery such as RhoA in regulating GBM cell mechanosensing and CSC emergence. Notably, Piezo1 were found to be co-localized with FAK within individual GBM cells experiencing mechanical constraints, which is consistent with a previous study that Piezo1 has important functions in focal adhesion dynamics in cells [59]. Higher expression of Piezo1 suggests enhanced focal adhesions thus larger traction force generation in cells in the CSC concentrated regions of GBM colonies. We further identified a positive association between Piezo1 and RhoA - a Rho family of small GTPase that is involved in actin-myosin contractility and stress fiber formation [63]. Upon mechanosensing by Piezo1, cells triggered its calcium signaling that activates RhoA, which then regulate contractility and reorganization of the actomyosin complex of the cytoskeleton, altering both intercellular and traction force generated by the cells.
In conclusion, using a 2D micropatterned multicellular model, we revealed that Piezo1, collaborating with focal adhesions, cadherins, and downstream cytoskeletal machinery, regulates mechanosensing of cancer cells to mechanical constraints in the GBM tumor microenvironment and guide the spatial patterning of in CSCs. These findings highlight Piezo1 as a potential mechano-therapeutic target to reduce tumorigenesis and drug resistance in GBM interventions.
Author contributions
Ngoc Luu: Conceptualization, Methodology, Investigation, Data curation, Formal analysis, Visualization, Writing - original draft. Shuhao Zhang: Investigation, Validation. Data curation, Formal analysis. Raymond H. W. Lam: Methodology; Supervision, Writing - review & editing. Weiqiang Chen: Conceptualization, Funding acquisition, Supervision, Writing - review & editing.
Ethical approval
This study does not contain any studies with human or animal subjects performed by any of the authors.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgement
This work was supported by the National Institutes of Health [R35GM133646].
Appendix A. Supplementary data
Supplementary data to this article can be found online at https://doi.org/10.1016/j.mbm.2023.100027.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}