
1. Introduction
According to the World Health Organization, 55 % of malignant tumors can be cured at present, of which the contributions of surgery, radiation therapy and chemotherapy are 27 %, 22 % and 6 % respectively [1]. Radiation therapy is used to treating a variety of cancers based on that cancer cells are more sensitive to radiation than normal cells. Various strategies such as radiosensitizers, radioprotective drugs, image-guided radiation therapy (IGRT), are used in the field of radiotherapy to maximize the dose of cancer cells and minimize the dose of normal cells [2,3]. Moreover, the precision of radiation dose delivery is rapidly improving by modern dynamic intensity modulated radiotherapy techniques such as intensity-modulated radiotherapy and volumetric modulated arc therapy. Nevertheless, acute and late injuries to radiation therapy continue to impact quality of life and even cause morbidity and mortality for cancer patients [4]. Patients almost invariably relapse and the recurred tumor has few treatment options. How to increase the lethality of cancer cells and improve the cancer patient prognosis is an urgent problem to be solved. Consequently, it is extremely significant to understand the functional consequences of radiotherapy and implement interventions that improve therapeutic effect and optimize patients' post-treatment quality of life.
Imaging modalities such as ultrasound elastography, magnetic resonance imaging (MRI) elastography, tissue diagnostic instrument (TDI) analysis and physical palpation often use tissue stiffness as a measure of breast tumorigenesis [5]. Matrix stiffness as one of the most important indicators exerts significant impacts on biological behaviors of cancer cells and associated cells such as growth, proliferation and apoptosis which are correlated with disease aggressiveness. On the one hand, the development and metastasis of tumors are often accompanied by excessive collagen deposition, cross-linking due to the increased expression of lysyl oxidase (LOX), remodeling, and the imbalance of matrix enzymes, resulting in increased stiffness in and around the tumor [6]. On the other hand, activated Rho-associated coiled-coil-containing protein kinase (ROCK) mediates actomyosin contraction and increased intracellular tension. Cells are drived to respond by altering ECM (increasing collagen deposition and the degree of crosslinking) to increase tissue stiffness and achieve intracellular and extracellular force balance [7]. In the past, researchers often focused on the change of matrix stiffness during tumor development and explored various treatment methods [8,9,10], while ignoring that the change in tumor matrix stiffness induced by radiotherapy also plays an important role in tumor regression and cure. An important reason for the latter is that ionizing radiation (IR) can also alter tissue density and rigidity.
High-energy X-rays cause apoptosis to rapidly proliferating cancer cells, and the radiation-induced activation and upregulation of proteases result in matrix proteolysis which can degrade the ECM [11,12]. Under normal circumstances, tumor tissues after therapy become soft significantly and decrease in volume such as cervical cancer and subcutaneous tumors [13], as shown in Table 1. Zeng research team [14] monitored the changes of stiffness in cervix and paracervical by shear wave elastography (SWE) to assist in evaluation of the efficacy of radiotherapy for cervical cancer. They found the elastic modulus of cervix decreased from 234.8 kPa to 167.6 kPa, indicating the recovery of cervical cancer. Zheng et al. [15] irradiated subcutaneous tumors in athymic nude mice model with a single 10 Gy dose. Mechanical changes in xenograft tumor samples were assessed by atomic force microscopy (AFM). IR ultimately resulted in a significant reduction in the stiffness of tumor tissues (22 kPa) compared to unirradiated tumors (46 kPa).
Table 1. Changes in the mechanical properties of different tissues before and after radiotherapy. SWE, Shear wave elastography. AFM, Atomic force microscopy. MFPs, Mammary fat pads. |
| Cancer types | Species | Mechanical performance index | Measurement methods | Irradiated site | Before/after radiotherapy | Ref. |
|---|---|---|---|---|---|---|
| Cervical cancer | Human | Elastic modulus | SWE | Cervical mass | 234.8 kPa/167.6 kPa | [10] |
| Subcutaneous tumors | Mice | Elastic modulus | AFM | Xenografted tumors | 46 kPa/22 kPa | [11] |
| Nasopharynx cancer | Human | Elastic modulus | Ultrasound | Neck | 12.8 ± 3.9 kPa/46.4-108.3 kPa | [12] |
| Breast cancer | Mice | Storage modulus | Rheometer | Pre-gel solution made from MFPs | low/high, no specific value | [13] |
| Breast cancer | Human | Shear wave velocity | Ultrasound with SWE | Pectoral muscle | low/high, no specific value | [14] |
| Head and neck carcinoma | Human | Shear modulus | Ultrasonog-raphy | Carotid arteries | 1.38 ± 0.98 kPa/3.10 ± 2.03 kPa | [17] |
| Pelvic cancers | Mice | Matrix stiffness | Mechanical stretcher | Bladder | low/high, no specific value | [18] |
However, high-energy X-rays may also have an important impact on normal tissues in the radiation field and the microenvironment that supports the growth of cancer cells [16]. The induction of normal cellular damage and local inflammation may further develop into fibrosis. ECM overproduction and remodeling result in an abnormal increase in the stiffness of normal tissues or tumor tissues. Significant differences of mechanical properties were also found between irradiated and non-irradiated breast tissues [17]. This may be related to changes in extracellular matrix (ECM) composition and mechanical properties. Irradiated skin closed to the arm by the side was significantly thicker and stiffer, and irradiated pectoral muscle was significantly stiffer [18]. When radiation therapy was extended to the upraclavicular and infraclavicular nodes, patients with breast cancer who received radiation had stiffer shoulders and were more likely to have poorer outcomes [19,20]. A recent study [21] also showed that the shear modulus was markedly different between irradiated (3.10 ± 2.03 kPa) and non-irradiated arteries (1.38 ± 0.98 kPa). This result suggests that the arterial shear modulus can be considered as a new biomarker of radiation-induced atherosclerosis in the common carotid artery. Additionally, Zwaans team [22] first proposed that changes in bladder mechanical properties were statistically apparent at 6 months post-irradiation in a radiation cystitis murine model received a single 40 Gy dose of X-rays. Radiation-induced subsequent changes in ECM content obviously increased the stress on bladder tissue when it filled with urine and subsequently stretches, causing significant changes mainly in voiding with a decrease in bladder capacity and a rapid increase in voided volume as the matrix stiffness increased (Table 1). Although IR is used as a standard treatment for a variety of malignant tumors, IR paradoxically promotes tumor recurrence and metastasis through the changes of matrix stiffness [23]. Increased tissue stiffness especially matrix stiffness caused by radiation has attracted more and more attention and has gradually become a new direction of intervention.
A comprehensive understanding of the biological regulatory mechanism of radiation on matrix stiffness and its impact on the effect of radiotherapy is beneficial for the development and screening of related drugs, optimizing clinical radiotherapy and ultimately improving the prognosis of patients. Hence, this review firstly introduces the cellular and molecular mechanisms of changes as well as influencing factors of matrix stiffness in different cancer tissues during radiotherapy. Then, the effects of radiation therapy-induced changes in matrix stiffness on cancer cells and associated cells are elucidated. Furthermore, relevant signal transduction mechanisms by which cells sense and respond to matrix stiffness are summarized. Finally, potential therapeutic strategies to improve the efficacy of radiotherapy based on matrix mechanics are discussed. In a word, matrix mechanics can be considered as a new monitoring biomarker of radiation-induced therapy resistance in common cancers.
2. Alteration of tissue matrix stiffness during radiotherapy
Normal tissue damage caused by radiation therapy is usually accompanied by acute or late toxicity, depending mostly on the tissue self-renewal time and its regulation process through the wound healing response. The acute phase occurs in the hours or days after radiation therapy and is caused by an inflammatory response, late radiation toxicity occurs after 9-12 months and involves an irreversible ECM remodeling [24]. Acute radiotherapy side effects may reduce the efficacy of radiotherapy or even make patients voluntarily give up radiotherapy for a long time. Besides, the perpetuation of radiation-induced fibrosis can promote decreased joint range of motion, lymphatic and vascular dysfunction as well as increased tissue stiffness [25].
In addition to direct damage to the proteins and DNA of cancer cells, radiation also indirectly increases reactive oxygen species (ROS), which can lead to other cellular damage, as shown in Fig. 1. These processes can lead to cell death or senescence after radiation [26], which can promote epithelial-mesenchymal transition (EMT), immunosuppression and tumorigenesis in neighboring cells, as well as re-entry into the cell cycle and activation of cancer stem cells, thereby promoting cancer cell survival [27,28]. Moreover, nitrogen species generated by IR can also lead to localized inflammation [29]. Clinically, patients may experience various adverse reactions during or after treatment which related to inflammatory response, such as fatigue, pain in breast cancer radiation therapy [30], vaginal rectal fistula in cervical cancer radiation therapy [31], urinary frequency in pelvic radiation therapy [22] and damaged esophageal mucosa caused by radiation esophagitis can also affect ingestion and acid reflux [32]. If the inflammatory response is maintained, inflammatory cells such as neutrophils, lymphocytes, and monocytes arrive at the site of injury which resultants M2 macrophages produce platelet-derived growth factor (PDGF), transforming growth factor-β1 (TGF-β1) and other pro-inflammatory factors, leading to recruit and stimulate the transdifferentiation of normal fibroblasts into cancer-associated fibroblasts (CAFs) [33]. CAFs secrete inflammatory mediators, excess collagen and other ECM components to remodel ECM [29]. In addition to change the amount of deposited collagen, the alignment of the collagen fibrils is also contributing significantly to the alteration of the matrix stiffness. The collagen matrix alignment in vivo can significantly improve the strength and stiffness of entire matrix due to its better structural order relative a matrix that is a disordered matrix. For example, the study found significant differences in elastic modulus between the aligned and random collagen type I scaffolds, with the aligned groups (567 ± 134 kPa) having significantly greater elastic modulus than random groups (349 ± 85.5 kPa). Besides the variation of orientation of collagen fibrils during the pathological progression of tumors, the change of collagen organization was also found to affect matrix mechanics [34]. Thickened and organized collagen fiber bundles are present in stiff tumor microenvironment. Besides, radiation promotes the secretion of LOX by cancer cells in a hypoxia-dependent manner which may lead to further cross-linking of collagen. These factors together result in the changes in ECM stiffness (Fig. 1). Dense ECM and abnormal vessel formation which contribute to tumor hypoxia increase the number of hypoxic cells and the difficulty of treating surviving cells [26,35,36,37].
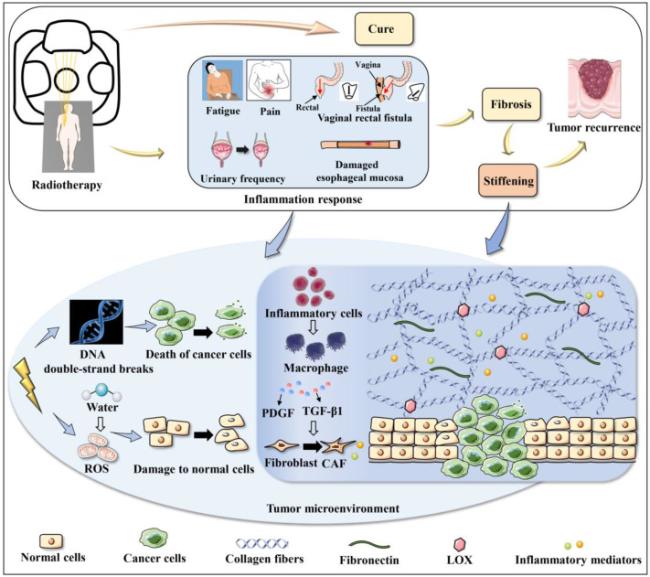
Fig. 1. Alteration of tumor matrix stiffness during radiotherapy. Some tumor patients are cured by radiotherapy, however, some patients may develop inflammation in tumors or surrounding normal tissues after radiotherapy, resulting in side effects of radiotherapy, such as fatigue, pain, urinary frequency, damaged esophageal mucosa, and vaginal rectal fistula. On the one hand, ionizing radiation breaks the DNA double strands to kill cancer cells. On the other hand, it ionizes water molecules to generate ROS to damage normal cells. Changes in the tumor microenvironment recruit inflammatory cells, secreting PDGF and TGF-β1 to stimulate the transdifferentiation of fibroblasts to CAFs, and the remodeling of ECM leads to fibrosis. The progression of fibrosis results in the increased ECM stiffness, associated with tumor recurrence. ROS, Reactive oxygen species. PDGF, Platelet-derived growth factor. TGF-β1, Transforming growth factor-β1. CAFs, Cancer-associated fibroblasts. ECM, Extracellular matrix. LOX, Lysyl oxidase. |
3. Clinical causes of tumor matrix stiffening during radiotherapy
Many clinical factors take effect on the changes of radiation on cancer cells and the tumor environment during radiotherapy individually or together. Total dose, fractional dose/fractional times of radiotherapy as well as dose distribution of radiotherapy are clinical factors that should be considered to cause changes in matrix mechanics of tumor tissues during radiotherapy. Understanding the differential effects of these factors can have implications for choice of combination therapies.
3.1. Total dose of radiotherapy
Kotova team [38] tracked the damaging effects of IR on the collagen structures by applying AFM. AFM imaging detected thick bundles of oriented collagen fibers with the quasi parallel fibrils inside the fibers both in the experimental animals’ bladder and rectum for a month post-irradiation in the dose of 2 Gy. The very early (a day post-irradiation) effects of increasing radiation doses (2, 8, 10, 12, and 22 Gy) were also investigated. Noticeable radiation damage increased proportionally with radiation dose. Especially at the 22 Gy dose, ECM collagen network of the normal bladder and rectal was restructured into the fibrotic-type collagen architecture. Similarly, Blockhuys research team [39] also observed a radiation dose-dependent increase of collagen type I reorganization by irradiated breast cancer cells in the stressed matrix contraction assay. The radiation dose varies from 0 Gy to 8 Gy, and the contraction index of collagen type I linearly varies from 10 % to 60 %. However, Miller and colleagues [40] investigated the effects of IR on an in vitro collagen matrix by using doses of 40 Gy and 63 Gy, respectively. The data indicates that IR can reduce the stiffness of collagen by breaking the connection between collagen fibers without significant changes in collagen fiber structure as the increased dose. This seemingly paradoxical phenomenon can be explained by the fact that the former is actually caused by radiation-induced destruction of tumor stroma and excessive damage to normal tissue. In the absence of these destruction conditions, matrix stiffness tends to decrease with increasing total dose. Radiation therapy doses cause varying degrees of collagen content changes that also impact chemotherapeutic drug delivery. Appelbe research team [41] exploited image-guided X-irradiation to augment nanoparticle accumulation in tumors. A single 5 Gy dose of radiation can markedly enhance the delivery of macromolecules and nanoparticles. However, the benefit of higher radiation doses (15 Gy) is limited due to increased collagen type I which severely damages blood vessels and reduces blood perfusion.
3.2. Fractional dose/fractional times of radiotherapy
The standard fractionation schedule for radiotherapy is 1.8-2.0 Gy per day, 5 days per week, which re-oxygenates tumor cells and redistributes their cell cycle to more sensitive phases, reducing tumor repopulation during treatment. Meanwhile, it allows for damaged normal cells to recover before additional doses are given. In clinical practice, the fractional dose of conventional radiotherapy for breast cancer is generally 1.8-2.5 Gy, the fractionated dose of hyperfractionated radiotherapy is < 1.8 Gy, and the fractionated dose of hypofractionated radiotherapy is 5-12 Gy [42]. Based on that, Qayyum and Insana [43] combined 3D collagen matrices, fibroblasts, and TGF-β1 to simulate the breast matrix microenvironment and study the effect of different fractionated doses of radiotherapy (90 cGy, 180 cGy, and 360 cGy) on collagen stiffness and fibroblast activation. They found that increased fractionated dose inhibited the formation of reactive stroma necessary for tumor progression and alleviated radiation-induced stroma stiffening, while decreased fractionated dose promoted fibroblast activation and possibly increased stroma stiffening (shear wave velocity: control-about 3.2 N/m, 90 cCy-about 1.7 N/m, 180 cCy-about 1.4 N/m). In another in vitro model mimicking the mammary interstitium after breast tumor resection [44], high fractionated dose of radiation inhibited fibroblast differentiation to reduce the oncogenic microenvironment more effectively. Nevertheless, it still appear to result in increased ECM stiffness (shear wave velocity: 90 cCy-about 1.2 N/m; 180 cCy-about 1.25 N/m; 360 cCy-about 2 N/m), which may stimulate the growth of cancer cells, indicating that matrix stiffness is an important factor in regulating the radiation sensitivity of cancer cells and associated cells [44].
3.3. Dose distribution of radiotherapy
High-precision radiation therapy technologies reduce the amount of normal tissue that receives a high dose of radiation but at the cost of large volumes of tissue receiving a low dose. It forces doctors and physicists to choose which organs or parts of the substructure of organ pairs should be preserved [45]. Dose distribution of radiotherapy is closely related to tumor tissue inflammatory response. Serious complications may occur if too much attention is paid to the target dose and the protection of organs at risk is neglected. Deterioration of the inflammatory environment is highly likely to cause changes in matrix stiffness [34,46]. In addition, the criteria based on biomechanical quality assurance (QA) is used in the radiotherapy deformable image registration (DIR) algorithm. The proposed biomechanical criteria imply the calculation of normal and shear mechanical stress and then compared to plausible physiological limits, providing the anatomical plausibility of the estimated deformations [47]. The criteria utilize the specific biomechanical properties of observed anatomy to provide a fast and objective physical interpretation of the estimated deformation, suitable for applications with low latency requirements such as adaptive IGRT.
4. Effects of matrix stiffness on the efficacy of radiotherapy
Matrix stiffness is not only an important outcome of tumor radiation therapy, but also an important regulator of the efficacy of radiation therapy. It regulates the ability of cells to develop forces through their contractile actomyosin cytoskeleton and provide feedback on the surrounding microenvironment. Altered matrix stiffness may trigger different cellular responses, which are closely related to tumor regression during treatment and tumor recurrence at the prognostic stage of treatment.
4.1. Contribute to the sensitivity of cancer cells and associated cells during radiotherapy
Radiation-induced changes in matrix stiffness of tumors can regulate radiation sensitivity of cancer cells and normal cells, which is extremely important to consider because it can inadvertently affect treatment response. Next, the biological behaviors of cancer cells, cancer stem cells, immune cells, and epithelial-like cells in 2D substrates or 3D matrices with different stiffness are elaborated.
4.1.1. Cancer cells
Increased matrix stiffness can promote damaged or senescent cancer cells to proliferate, migrate, invasion and even alter radiosensitivity [48]. Thus, it is logical to infer that a stiff tumor microenvironment is more likely to facilitate resistance to therapy [49] (Fig. 2). Glioblastoma multiforme (GBM) is the most aggressive type of brain cancer and cannot be completely removed by surgery so that recurrence is inevitable. Due to the highly heterogeneous and various matrix compositions, GBM tumors often experience treatment failure and are prone to develop drug resistance [50]. Marhuenda and colleagues [51] designed 3D-ex-polyacrylonitrile nanofibers scaffolds (NFS) with adjustable stiffness to study the effect of matrix stiffness on the migration behavior of glioblastomas stem-like cells (GSCs). They found the stiffness of 166 kPa is sufficient to trigger migration, resist to apoptosis and produce of ECM components. Experimental result also demonstrates that cervical squamous carcinoma SiHa cells grown on hydrogels with different stiffness (0.5, 5, and 25 kPa) showed different morphology, proliferation rate and radiosensitivity. Cells grown on substrates with the stiffness of 25 kPa displayed larger size, more mature cytoskeleton assembly. Furthermore, cells on soft substrates (0.5 kPa) grew more slowly and were more sensitive to radiation, possibly via altered apoptosis-related protein expression [31].
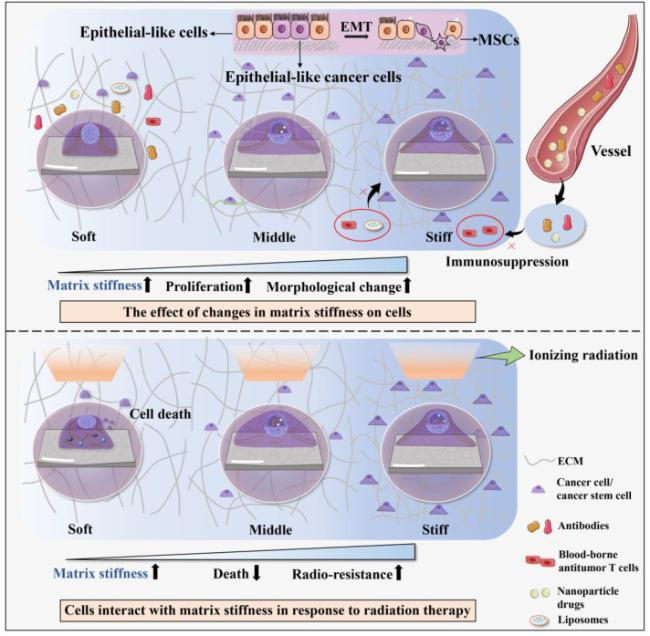
Fig. 2. Contribute to the sensitivity of cancer cells and associated cells during radiotherapy. The stiffness of irradiated ECM is increased, and the morphology of cancer cells or cancer stem cells gradually change from round to spread. Moreover, the proliferation ability of cells on stiffer matrices is greater than that on softer matrices. In the presence of altered tumor mechanical microenvironment, a stiffer microenvironment is more prone to immunosuppression and EMT. After cells are irradiated, the behaviors of cells are different with different matrix stiffness. Softer matrices are more likely promote cellular radiosensitivity, while stiffer matrices can inhibit cellular radiosensitivity. EMT, Epithelial-mesenchymal transition. MSCs, Mesenchymal stem cells. |
Moreover, cancer cells can also interact with matrix stiffness in response to irradiation dose. Panzetta and colleagues [52] evaluated adhesion and migration properties of MCF10A and MDA-MB-231 cell lines, over a range of pathophysiological stiffness (1-13 kPa) before and after the exposure to two different X-ray doses (2 and 10 Gy, photon beams). The matrix stiffness of normal tissue was radioprotective against cell motility and invasion in both cell lines. The higher matrix stiffness (supraphysiological matrix stiffness) instead promoted tumor cell motility (Fig. 2). Nevertheless, the impact was completely eliminated by the cells irradiated with the higher dose (10 Gy) in a long time. This result suggests that changes in cancer cell behaviors caused by changes in matrix stiffness during radiation therapy can be restored by adjusting the irradiation dose. In contrast, not all experimental results are consistent. Lacombe et al. [53] studied the effect of matrix stiffness on the radiation response of lung cancer cells by using polyacrylamide (PA) collagen-coated hydrogel model. Lung cancer cells seeded on both soft (0.7 kPa) and stiff (70 kPa) hydrogels were irradiated, and the level of DNA damages were compared. Total amount of DNA damages was higher in cells grown on a stiff ECM at 1 h post-irradiation than cells seeded on a soft matrix, partially by remodeling chromatin that could expose DNA to radiation-induced genotoxic agents such as ROS. Similarly, mammary epithelial cells and tumor cells assembled into organoid-like structures in soft (140 Pa) ECMs can efficiently engage NF-κB signaling to resist apoptosis induction. Conversely, cells interacting with a stiff (5 kPa) ECM showed reduced NF-κB signaling and increased c-Jun NH2-terminal kinase (JNK) activity that sensitized them to several apoptosis-inducing agents [54]. Compared to monolayer culture studies, recapitulating the 3D tissue-like structures of in vivo tumor tissue by organoid was faithfully reproduced in mouse models and patient samples. It is obvious that the establishment of the experimental platform also significantly affects the experimental results. Besides, this seemingly paradoxical phenomenon may also be associated with different types of tumors and setting of platform stiffness range. Diverse cells have different sensitivities to matrix stiffness as well as different stiffness setting criteria, leading to the differences in experimental results. Therefore, it should be more accurate to study the cellular response to matrix stiffness by setting the same matrix stiffness classification.
4.1.2. Cancer stem cells
Cancer stem cells (CSCs) are responsible for malignant progression and clinical data shows that CSCs can survive during radiation therapy [55]. Additionally, an increasing body of studies have shown that matrix stiffness has a significant effect on the properties of cancer stem cells. Liver cancer cells with stemness properties (LCSCs), a type of cancer stem cells, comprise a small portion of hepatocellular carcinoma (HCC) cells. LCSCs have the characteristics of great self-renewal and metastatic potential and are insensitive to drugs and radiotherapy, playing a key role in the recurrence and metastasis of HCC. Tian and colleagues [56] utilized mechanically tunable PA hydrogels to simulate different stiffness of the liver tissue matrix in various stages. The results showed that HCC cells were small and round in the soft matrix but better spread on the stiff matrix, suggesting that less biomechanical stimulation is beneficial for maintaining cancer cell stemness (Fig. 2). This is of great significance for effectively controlling tumor matrix stiffness to prevent tumor recurrence during radiotherapy or in the prognostic stage. However, this relationship may not apply to all tumor types and stages. You et al. [57] proposed that HCC cells maintain high stemness on gels up to 16 kPa via activating the integrin β1/Akt/mTOR/SOX2 signaling pathway, resulting in high expression of CD133 and epithelial cell adhesion molecule (EpCAM). In addition, integrin-linked kinase (ILK) is an important mechanotransducer of mammary CSCs. The expression of CSC marker depends on stiffness and hypoxia. Stiff microenvironments promote tumor formation and metastasis in ovo through the modulation of ILK, whereas ectopic expression of ILK also stimulates CSC development even under softer or normoxic conditions [58].
4.1.3. Immune cells
Radiation therapy can enhance tumor lethality by increasing the number of tumor-infiltrating immune cells and the expression of neoantigens. It has been reported that radiotherapy can induce immunosuppression in addition to anti-tumor immune promotion. Tumor stiffening is due to increased deposition of interstitial collagen, hyaluronic acid, elastin, and fibronectin, as well as increased numbers of benign and malignant cells. Mechanical forces mediated by these structural components can constitute a physical barrier that impedes the entry and movement of blood-borne antitumor T cells, antibodies, liposomes, and nanoparticle drugs. It greatly affects immune surveillance and immunotherapy response [59] (Fig. 2). Therefore, matrix stiffness may become a problem for pursuing tumor immunotherapy following radiotherapy. Dose and fractionation have also been demonstrated to play a role in the balance of immune priming and immunosuppression after radiation therapy [60]. Due to the double-edged sword effect of radiation therapy on immunomodulation, precise dosing regimens and combination therapy with radiation therapy must be carefully considered to avoid unnecessary immunosuppressive effects [61].
4.1.4. Epithelial-like cells
EMT can be induced through the activation of EMT signaling pathway and ECM remodeling in tumor microenvironment [62]. Epithelial cells may undergo a mesenchymal transition in a rigid environment (Fig. 2). Fibrotic stiffness elements found in pancreatic cancer tissue, including increased vimentin expression, decreased E-cadherin expression, and nuclear localization of β-catenin, promoted EMT development [63]. Recently, EMT was also found to contribute to radiation resistance mechanisms in solid tumors. As epithelial cancer cells undergo EMT, they gain stemness, invasiveness, motility, angiogenic and metastatic ability. Gomez et al. [64] demonstrated non-small cell lung cancer cells that survived ionizing radiation treatment display cancer stem cell and EMT phenotypes. Matrix stiffness has also been shown to alter the effects of TGF-β signaling, from promoting apoptosis to inducing EMT in epithelial cells. Decreasing stiffness increased TGF-β1-induced apoptosis of epithelial-like cancer cells, whereas increasing stiffness resulted in EMT in response to TGF-β1 [65].
4.2. Contribute to the tumor recurrence at the prognostic stage of radiotherapy
Changes in matrix stiffness during radiotherapy induce the changes of cancer cell radiosensitivity and the antitumor behavior of associated cells. This series of responses may enhance tumor resistance to radiotherapy, and increase the probability of tumor recurrence in the prognostic stage [66,67]. The inflammatory processes that arise immediately after radiotherapy are not the same as those that appear decades later. Early biological events cause acute tissue effects, which are normally resolved within three months of completing treatment. They also may result in more protracted biological effects which can manifest in tissues as late fibrotic states or secondary malignancies [68]. In addition to collagen content, the alignment and orientation of collagen fibers appear to play a key role in the changes of matrix stiffness [34], and aligned collagen is shown to be a prognostic feature of human breast cancer survival. Aligned collagen can predict poor prognosis in breast cancer patients, possibly because it promotes invasion by providing tracks for cells to migrate out of the tumor [69,70]. It is necessary to pay more attention to the improvement of prognosis while increasing the radiosensitivity of cancer cells by matrix stiffness.
5. Mechanical signal transduction mechanism in radiotherapy
The signals pathways that mediate cells to sense and respond to the changes of matrix stiffness have been intensively studied [9,71]. In order to increase the anticancer effects of tumor therapy, it is vital to understand the underlying mechanisms of matrix stiffness promoting tumor formation. Next, as shown in Fig. 3, signal transduction pathways involved in ionizing radiation-induced changes in matrix stiffness to trigger changes in cell behaviors are summarized.
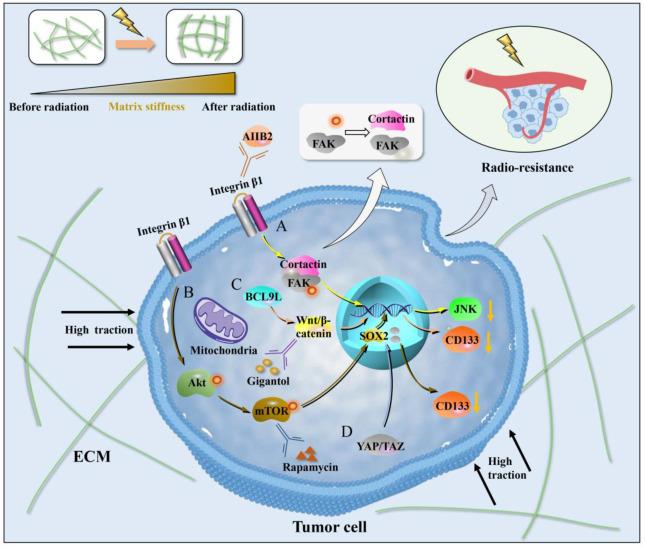
Fig. 3. Mechanical signal transduction mechanism in radiotherapy. (A) Integrin β1/FAK/cortactin signaling pathway. Integrin β1 transmits ECM mechanical signals to intracellular FAK/cortactin, and phosphorylated FAK transmits signals to the nucleus, resulting in the transcription and expression of related genes. After inhibition of integrin β1 by AIIB2, dephosphorylation of FAK leads to the dissociation of FAK/cortactin protein complex. The expression of JNK signaling decreases, and cells are radiosensitive. (B) Akt/mTOR/Sox2 signaling pathway. Increased matrix stiffness can upregulate the phosphorylation levels of Akt and mTOR and promote the expression of Sox2 gene through integrin β1. MTOR inhibitor rapamycin suppress mTOR phosphorylation levels and SOX2 expression levels in HCC cells grown on high stiffness substrates, and significantly suppress the expression of CD133. (C) BCL9L/Wnt/β-catenin signaling pathway. Wnt/β-catenin expression is higher in the stiff matrixes than in the soft and moderately stiff matrixes, and silencing of BCL9L can reverse this phenomenon. Inhibition of Wnt/β-catenin signaling pathway using gigantol can significantly down-regulate the expression of CD133. (D) YAP/TAZ signaling pathway. Elevated matrix stiffness can induce massive nuclear transcription and activity of YAP/TAZ. FAK, Focal adhesion kinase. AIIB2, Anti-integrin β1 antibody. JNK, c-Jun NH2-terminal kinase. YAP, Yes-associated protein. TAZ, Transcriptional coactivator with PDZ-binding motif. |
5.1. Integrin β1/FAK/cortactin signaling pathway
The integrin-mediated signaling pathways necessary for tumor development and radiation response are significantly altered by ECM reorganization in head and neck cancers and breast cancers. Integrins and their downstream focal adhesion proteins (FAPs) are known as mechano-sensors and mechanotransducers that sense and transduce mechanical signals into chemical signals. Integrin β1 is the most common subchain of integrin heterodimers. Multiple studies have shown that integrin β1 is widely overexpressed in tumors such as lung, breast and plays an important role in their survival and metastatic potential [72,73]. Integrin β1/focal adhesion kinase (FAK)/cortactin signaling pathway has been proved to be an important signaling pathway for radiotherapy resistance in head and neck cancer (Fig. 3A). Integrin β1 may induce radio-resistance via affecting DNA repair and Yes-associated protein 1 (YAP1)-induced EMT, and may be an attractive therapeutic target for overcoming radio-resistance in non-small cell lung cancer (NSCLC) cells [74]. FAPs also essentially contribute to radiotherapy resistance, and the possibility of targeting these proteins could be developed as a therapeutic option in combination with radiotherapy and chemotherapy [75]. Dephosphorylation of FAK following inhibition of integrin β1 by anti-integrin β1 antibody (AIIB2) leads to dissociation of the FAK/cortactin protein complex. This in turn inhibits JNK signaling and induces rounding of cancer cells, resulting in radiosensitization [76].
5.2. Akt/mTOR/Sox2 signaling pathway
Substrate stiffness can cause changes in the characteristics of tumor stem cells, which are related to the activation or inactivation of some signaling pathways, mainly involving integrin β1/Akt/mTOR/Sox2 signaling pathway [77] (Fig. 3B). You et al. [57] found that increased matrix stiffness can upregulate the phosphorylation levels of Akt and mTOR and promote the expression of Sox2 gene. Conversely, knockdown of integrin β1 partially reversed the phosphorylation levels of Akt and mTOR as well as the expression of SOX2, and attenuated high stiffness-mediated stemness characteristics in HCC cells, suggesting that integrin β1 may transmit higher stiffness signals to HCC cells and activate the mTOR signaling pathway. Furthermore, mTOR inhibitor rapamycin suppressed mTOR phosphorylation levels and SOX2 expression levels in HCC cells grown on high stiffness substrates, and significantly suppressed their stemness properties. Therefore, matrix stiffness can participate in the regulation of HCC stemness by activating integrin β1/Akt/mTOR/SOX2 signaling pathway.
5.3. BCL9L/Wnt/β-catenin signaling pathway
Wnt signaling is a critical determinant of CSC pluripotency and self-renewal, and BCL9 and BCL9L are transcriptional co-activators within the Wnt enhanceosome [78]. The investigation of the underlying mechanism indicated that higher matrix stiffness increased glioma stemness resulting in sustained tumor growth by activating the BCL9L/Wnt/β-catenin signaling pathway [79] (Fig. 3C). In this phenomenon, the expression of BCL9L was obviously up-regulated. Similarly, researchers observed that Wnt/β-catenin expression was higher in the stiff (16 kPa) polyacrylamide hydrogels than in the soft (2 kPa) and moderately stiff (8 kPa) groups, but silencing of BCL9L reversed this phenomenon. Inhibition of Wnt/β-catenin signaling pathway using gigantol can down-regulate the expression of CD133 and sustainably improve the anticancer effect of radiotherapy in mice with subcutaneous glioma. The results indicate that intervention of drugs during radiotherapy can greatly improve the effect of tumor treatment compared with radiotherapy alone [79].
5.4. YAP/TAZ signaling pathway
A wealth of experimental evidence now supports that YAP mediates radiotherapy resistance [80]. YAP and its paralog tran scriptional co-activator with a PDZ-binding domain (TAZ) are important mechanistic transcriptional regulators, playing an important role in modulating cell proliferation, apoptosis, invasion, and migration. It is clearly demonstrated that elevated matrix stiffness induces massive nuclear transcription and activity of YAP/TAZ [81,82] (Fig. 3D). Increased nuclear localization of YAP/TAZ and higher transcriptional activities of YAP/TAZ target genes have also been observed in therapy resistant tumors [83]. In addition, YAP overexpression protected while YAP knockdown sensitized urothelial cell carcinoma (UCC) cells to radiation effects by increasing the accumulation of DNA damage and apoptosis [84]. The research found that high levels of YAP function in the stroma are sustained through positive feedback between CAF-driven matrix stiffening and mechano-transduction leading to YAP. YAP regulates the expression of multiple cytoskeletal regulators in CAFs, including ANLN and DIAPH3, and controls the protein levels of MYL9/MLC2. Matrix stiffening further enhances YAP activation, thereby establishing a feed-forward self-reinforcing loop that helps maintain the phenotype of CAFs and further promotes tumor development [85]. Therefore, modulation of YAP/TAZ activity in cancer cells can clearly influence sensitivities to anti-cancer therapies.
6. Strategies to enhance the efficacy of radiotherapy based on matrix mechanics
As an important physical property of ECM, matrix mechanics is not only an important pathological consequence of ionizing radiation but also hinders radiotherapy. Therefore, from the perspective of matrix mechanics, effective methods to treat tumors can be explored shown in Fig. 4.
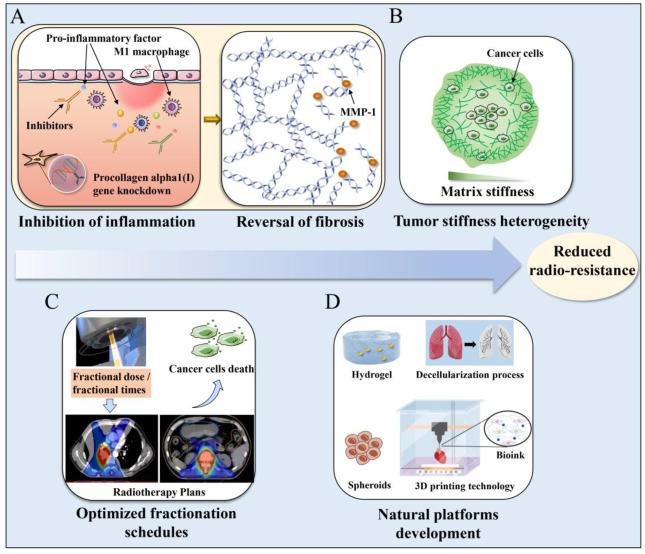
Fig. 4. Strategies to enhance the efficacy of radiotherapy based on matrix mechanics. (A) Targeting pro-inflammatory factor, receptor kinases, or matrix enzymes and reversing radiation-induced fibrosis. (B) Pay more attention to tumor stiffness heterogeneity. (C) Optimize total dose and fractionation schedules. (D) Development of biomimetic in vitro models. MMP-1, Matrix metalloproteinase-1. |
6.1. Targeting inflammatory response and reversing radiation-induced fibrosis
Tumor irradiation can induce a wound healing response characterized by inflammation, CAF modulation, ECM remodelling and ultimately results in increased matrix stiffness. Experimental results demonstrated that hard substrate (29 KPa) induces M1 macrophages and severe inflammatory response [86], which further promotes an increase in matrix stiffness and the resulting negative feedback leads to tumor recurrence. Targeting the initial inflammatory response may counteract attempts to boost the immune-mediated anti-tumor response following radiotherapy (Fig. 4A). Therefore, reducing ECM remodelling by inhibiting pro-inflammatory factor, such as TGF-β, NF-κB, COX-2 and TNF-α, receptor kinases, or matrix enzymes may be more effective in preventing the post-irradiation stiffening of tumor microenvironment that could facilitate tumor metastasis [87]. In addition, procollagen alpha1 (I) gene knockdown mediated by shRNA also downregulated procollagens type III, IV and VI and other inflammation related parameters [88]. Collagen type I, as a modulator of other collagens and a promoter of chronic inflammation, may also be an extremely important target during radiotherapy-induced tumor eradication.
Several approaches against radiation-induced fibrosis have also been evaluated clinically, but the effectiveness of these interventions for the treatment of radiation-induced fibrosis in cancer patients could not be determined [89]. The increase in matrix stiffness caused by radiation therapy may reduce the radiosensitivity of cancer cells, and the effect of radiation therapy with other drugs may be better during radiation therapy. Matrix metalloproteinase (MMP)-1 enzymatically degrades matrix proteins to soft the matrix (Fig. 4B). In Liang team's research [90], hepatoma cells were cultured in collagen-based gels with an elastic modulus of approximately 4 kPa. The elastic modulus of gel was decreased from 4 to 0.5 kPa by subsequent exposure to MMP-1. Hepatoma cells respond to MMP-1 by actively proliferating, downregulating E-cadherin, and losing their ability to detoxify. The resulting spheroids were more sensitive to radiation than spheroids cultured in stiffer gels without MMP-1. Radiation-induced fibrosis can be reversed by interventional means targeting the fibrotic ECM to reduce matrix stiffness.
6.2. Pay more attention to tumor stiffness heterogeneity
Tumors and their microenvironments are highly heterogeneous. Therefore, the mechanics of different regions within the tumor may also be different [91]. Anatomic imaging has become a critical component in the radiotherapy treatment planning and the treatment response assessment of sarcomas. Pepin et al. [92] measured the stiffness of a pelvic myxoid liposarcoma using magnetic resonance elastography (MRE) and found the stiffness heterogeneity within the tumor [93]. Correlative stiffness maps of different intratumor locations in breast tumor biopsies showed that stiffness increased from core to periphery [94] (Fig. 4B). Therefore, differences in local ECM stiffness may cause different mechanical responses of cancer cells, especially during radiation therapy, where changes in cell behavior also imply changes in radiation resistance. It is imperative to consider the heterogeneity of oncogenic ECM rather than isolated characteristics such as increased stiffness. Studying how this biomechanical heterogeneity affects the biological behavior of cancer cells has implications for the development of appropriate therapeutic schedule. Non-uniform radiotherapy fractionation schemes may be considerably more effective, that is, for the first stage, the radiosensitivity of tumor cells is increased, and for the second stage, the dose in areas of greater matrix stiffness can be increased. Tumor stiffness heterogeneity can be used with currently employed diagnostic methods to better predict treatment response and prognosis.
6.3. Optimized total dose and fractionation schedules
Clinicians prescribe appropriate doses of radiation therapy according to the radiobiological effects of cancer cells and individual circumstance, but ignore the possible impact of matrix stiffness on cancer prognosis. This could have important impacts on physicists in planning radiation therapy for cancer patients. Optimizing the radiotherapy prescription conditions from the perspective of tumor matrix mechanical state can provide a new entry point for improving the radiotherapy effect of tumors. Conventionally, delivery of doses of 50-70 Gy using multiple fractions of 1.8-2 Gy per fraction is the first choice for the optimal ratio of tumor treatment. And the advent of hypofractionated and macrofractionated delivery schemes in recent years has been used to treat many tumor types. Radiation can elicit immune effects in the tumor microenvironment, but the optimal dose and fractionation schedule required to induce immune priming are also unclear. Therefore, the immune effect is a double-edged sword. Different total dose and fractionation schedule also have an impact on physical planning (Fig. 4C). If an OAR of the same patient in two different treatment plans is irradiated much more in one plan than the other, each plan would have its optimal fraction size. Clinically, radiation treatment schedules can be optimized on a patient-by patient basis through radiobiological analysis. Alternative schedules which meet the objectives and constraints of radiotherapy prescription can be provided for physicians through the optimization process [95]. How to regulate the matrix mechanical state of tumor by optimizing the fractionated dose of radiotherapy and better induce cancer cell apoptosis or promote immune response still need further research.
6.4. Development of biomimetic in vitro models
At present, most studies mainly focus on 2D platforms or quasi-3D platforms which lack the interaction between cells and the natural microenvironment, and cannot reflect the regulation effect of changes in matrix stiffness induced by in vivo radiation therapy on cancer cells [44,96]. The development of a 3D biomimetic platform closer to the natural microenvironment will be more accurate for studying the effect of matrix stiffness on cancer cells during radiotherapy and its specific mechanisms (Fig. 4F). Certain new tumor-targeting techniques for radiation therapy such as MRI-guided radiation therapy and proton therapy techniques, are emerging. However, the mechanobiological effectiveness of these emerging technologies as compared to established photon-based radiotherapy is understudied. For example, for MRI-guided radiotherapy, the new use of static magnetic fields (SMFs) during radiation delivery needs to be thoroughly investigated. Tissue engineering including organoids, spheroids, hydrogels, developing as a promising biomimetic approach, can generate 3D models that capture structural, biophysical, biochemical, and biomechanical features. Therefore, it facilitates to more realistic treatment response studies for further clinical applications [91]. The decellularization process removes cellular and nuclear material, while preserving the important components of ECM and the structural and functional characteristics of the original microvascular network. In recent years, decellularized matrix has been widely used in tissue repair and the establishment of bio-platforms [97,98,99,100]. 3D printing technology can shape materials into desired shapes, and has been widely used in medical, radiation therapy and other fields in recent years [101]. Bioink is composed of water-rich molecules, a variety of cells and chemicals that promote cell communication and growth. It is used for bioprinting to reproduce the structure and function of tissues and organs. When consider the biophysical properties of the matrix, the combination of 3D printing technology and decellularization technology can provide natural and powerful conditions for further study of radiation therapy-induced changes in tumor matrix stiffness leading to changes in cell behavior in the future [102].
7. Conclusions and perspectives
The great lethality of high-energy rays on malignant tumors makes radiotherapy more and more favored by patients. However, due to the adverse reactions caused by radiation, medical personnel and researchers are gradually paying more attention to the prognosis of tumor patients. Understanding the mechanism of this adverse reaction may help to inform choice and improve response to combination therapies. Malignant cancer cells with genetic abnormalities are closely associated with a tumor stroma composed of diverse cellular and acellular entities, including CAFs, ECM components, immune cells, stem cells, and secreted bioactive molecules. Matrix stiffness, as an important physical feature of ECM, profoundly affects their highly dynamic interactions and controls tumor initiation, progression, invasion, metastasis, and therapy resistance. Appropriate matrix stiffness can enhance the sensitivity of cancer cells to radiation and increase the lethality of cancer cells [103]. Although many studies have preliminarily demonstrated that radiation therapy induces changes in matrix stiffness and it has profound effects on a wide range of cells, future work is needed to discern the effect of radiation therapy on each component of ECM and the overall change in tumor mechanical properties.
Moreover, application of non-invasive detection methods will greatly promote the detection of changes in matrix stiffness, speed up disease diagnosis and make judgments on the direction of treatment. In addition to traditional tissue stiffness detection methods such as ultrasound elastography, MRI elastography, the combination of computer science and biomechanics may become a new direction for future research, innovation, and development. Artificial intelligence and machine learning can be leveraged in the future to gain insights into radiomics based on stiffness signatures to inform future risk stratification [104]. By building a deep learning model to quickly learn mechanical features from the data set, predicting changes in tissue stiffness may provide a favorable reference for diagnosing disease development in patients.
In short, tissue mechanics, especially matrix stiffness, has great clinical value in the field of tumor radiation therapy, but the premise is that the underlying mechanism of its radiation-induced changes and further effects on cell behavior must be clearly studied. Translating these insights into clinical and therapeutic interventions may guide the design of safer and more effective cancer treatments.
Ethics declarations
This study does not contain any studies with human or animal subjects performed by any of the authors.
Consent for publication
Not applicable.
Data availability
Data sharing is not applicable to this article as no datasets were generated or analyzed in this study.
Credit authorship contribution statement
Yaxin Deng: Conceptualization, Writing - original draft, Writing - review & editing. Hong Deng and Guobao Chen: Literature collection, Figure preparation. Jiali Xiao: Supervision, Writing - review & editing. All authors read and approved the final manuscript.
Declaration of interests
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgements
This work was supported by the Bureau of Science & Technology Nanchong City [grant number No. 22SXQT0323].

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}