
1. Introduction
The gap between basic and clinical research has consolidated in recent years the view that two-dimensional cultures cannot efficiently recapitulate the natural environment of cells and tissues. This is mainly because the cells in vivo are organized and function in three-dimensional niches with cellular and acellular cues. This established knowledge, combined with the opportunity offered by the blooming and the evolution of the scaffold fabrication techniques, allowed the translational scientists to refocus their interest from the conventional 2D cultures to the 3D artificial microenvironments [1]. Hence, scientists from different fields joined forces towards the creation of 3D structures for culturing cells and tissues due to their evident advantages in providing more physiologically relevant information and more predictive data for in vivo tests [2,3,4,5]. To this end and inspired by nature, several biomimetic patterned surfaces with regularly or randomly arranged nano- and micro-structures protruding from a flat surface have been designed as suitable platforms for cell interfacing. Different geometries (nanowires, nanopillars, mushrooms, nanocavities) [6,7], structural densities, materials and fabrication technologies, as well as diverse cellular models, have been investigated. The growing interest in micro- and nanostructures as cell substrates originates from a number of promising applications, ranging from tissue engineering and regenerative medicine to neuroscience, pharmacology and neural computation [8,9,10,11,12,13].
There are many reports showing that the patterning of a surface with high-aspect-ratio micro- and nano-structures, although it may not provide a full 3D microenvironment can dramatically alter cell interactions with the material. Such textured surfaces having features on similar length scales to cellular and subcellular components can perturb the ability of a cell to sense and respond to microenvironmental cues, adapting its behavior. Cells are extremely sensitive to the topographical information of their environment, being able to react to objects as small as 5 nm [14,15]. The ability of cells to sense mechanical cues and translate them into a biological response by altering cell behavior, morphology and eventually gene expression, is taking place through a series of mechanisms that are collectively referred as mechanotransduction. In this context, defined surface topographies were shown to selectively guide cell responses. Surface roughness and specific surface features at subcellular scale are able to affect cell orientation, morphology, proliferation and differentiation [16,17,18]. Among them, differentiation and guidance are important in neural tissue engineering where controlling neurite extension, neuronal polarity and maturation process towards the formation in vitro of fully functional networks with defined connectivity is extremely challenging.
The unique properties exhibited by natural multifunctional surfaces are attributed to hierarchical structures in the micro-and nanometer length scale that often decorate these surfaces the mimicking of which can recapitulate better the in vivo microenviroment [19]. The creation of such natural-inspired surfaces, tailored for particular applications, requires a processing methodology that enables highly reproductive though delicate, and precise surface modification. Materials processing, using ultrafast lasers meets such requirements for fast and precise surface patterning, usually without the need for pre- or post-processing treatment [20,21,22,23].
Pheochromocytoma PC12 cells, is a typical cell model [24,25] that has been used extensively to study neuronal differentiation as upon exposure to nerve growth factor (NGF) morphologically and functionally mature sympathetic neurons are formed. It has been shown that PC12 cells are extremely sensitive to mechanical cues and respond to topographical features by modulating their differentiation and guidance accordingly. When cultured on micropatterned Si wafer, it has been demonstrated that PC12 cells oriented along the scaffold's grooves and the neurite outgrowth was enhanced compared to the flat surface [26]. Similarly, micro-nanostructured Pt or Si surfaces decorated with spikes with varying inter-spikes distance, enhanced PC12 differentiation [27]. Also, the neurite length of PC12 cells treated with NGF was found to be higher on patterned PDMS compared to the flat surface and more differences could be observed on the neurite length between isotropic and anisotropic microstructures [28].
In our previous work, we have used laser patterned Si substrates decorated with microcones as culture platforms for neuronal studies using the rat PC12 cell line [17]. In this study, PC12 cells were cultured on Si substrates comprising pseudoperiodic arrays of elliptically cross-sectioned microcones (MCs), fabricated using ultra-short pulsed laser processing of planar Si. It has been shown that the patterned surfaces exhibiting large distances among MCs (referred to as high roughness), did not support PC12 cell differentiation despite the presence of NGF. This observation is compelling as it implies that topographical cues could act superiorly to the chemical signals. In a similar study [29], the differentiation of neuronal cells (Neuro2A cell line) was inhibited on the high roughened Si substrate as well as on its polymeric replicas, despite the presence of differentiation medium which means that this phenomenon it is not restricted to the cell type nor to the type of material. In the current study, we attempt to elucidate the mechanisms underlying this intriguing cell behavior, via the investigation of key molecules participating in the mechanotransduction machinery, taking place during cell-surface interaction. Understanding neurite differentiation, outgrowth and orientation, in relation to topography of the extracellular environment, is important for the design of biomaterials that interface with the neuronal tissue and aim to reveal the biological requirement for neural network rewiring.
2. Materials and Methods
2.1. Antibodies and reagents
The following reagents and antibodies were used: DMEM 1X (41966-029, Gibco), 10X PBS (70011-044, Gibco), 10X PBS with Calcium and Magnesium chloride (D1283, Sigma), Horse serum heat inactivated (26050088 Thermoscientific LSG), Fetal Bovine Serum (F7524, SIGMA), Bovine Serum Albumin Fraction V (10735078001, Roche), 100 units/ml penicillin-streptomycin (PEN-STREP, Gibco, Thermo Fisher Scientific) Poly-d-lysine hydrobromide (P6407, SIGMA), Laminin from Engelbreth-Holm-Swarm murine sarcoma basement membrane (L2020, SIGMA), mouse Nerve Growth Factor 2.5S (N-100, Alomone Labs), mouse anti-Tubulin β3, 1/1000 (801213, Biolegend), mouse anti- Vinculin (clone 7F9) 1/400 (Part No. 90227, FAK100 Kit, Millipore), TRITC-Phalloidin 1/1000 (Part No. 90228, FAK100 kit, Millipore) rabbit anti-YAP clone D8H1X, 1/100 (14074, Cell Signaling Technology), rabbit anti-Phospho-Myosin Light Chain 2 (Ser19), 1/100 (3671, Cell Signaling Technology), DAPI (ProLong™ Gold Antifade Mountant with DAPI, ThermoScientific LSG).
2.2. Fabrication and characterization of microconically structured Si surfaces
The fabrication and the characterization of the patterned Si surfaces were performed as described previously [16,17]. Briefly, Single crystal n-type Silicon (1 0 0) wafers were subjected to laser irradiation in a vacuum chamber using as a laser source a regenerative amplified Ti:Sapphire (l ¼ 800 nm) delivering 150 fs pulses at a repetition rate of 1 kHz. After laser irradiation, an image-processing algorithm (ImageJ, National Institutes of Health, Bethesda, MD, USA) was implemented to determine the topological characteristics of the MCs, including height (h), base diameter (d), aspect ratio (A) and roughness ratio (r) from top, side-view and cross-sectional SEM images. The geometrical characteristics of the different substrates are seen in the following table (Table 1) adapted from [16,17].
| Type of roughness | Density, D ± STDEV (106/cm2) | Height, a ± STDEV (μm) | Base major axis, b ± STDEV (μm) | Base minor axis, b’ ± STDEV (μm) | Interspike distance, c ± STDEV (μm) |
|---|---|---|---|---|---|
| Low | 9.75 ± 1.54 | 1.26 ± 0.28 | 1.96 ± 0.35 | 1.11 ± 0.20 | 2.59 ± 0.72 |
| Medium | 5.01 ± 0.19 | 3.76 ± 0.42 | 3.57 ± 0.43 | 1.98 ± 0.34 | 4.71 ± 0.37 |
| High | 2.50 ± 0.26 | 8.63 ± 1.17 | 7.25 ± 0.72 | 3.78 ± 0.77 | 6.50 ± 0.84 |
2.3. Coating of surfaces
To enhance cell adhesion, the glass slides and the Si surfaces were all double coated with PDL and laminin. One day before seeding all culture surfaces were coated with 100 μg/ml PDL for 2 h at 37 °C. After incubation they washed 3 times with 1X PBS and they left to air dry. The following day, the PDL-coated samples were coated with 20 μg/ml laminin for 2 h at 37 °C. After the incubation, the laminin solution was removed and the cells were seeded immediately without washing.
2.4. Cell culture
The rat pheochromocytoma PC12 cell line (ATCC-LGC, Rockville, MD, USA) was maintained in DMEM supplemented with 10 % v/v HS and 5 % v/v FBS, at 5 % CO2, 37 °C. Only low passage number (P2-10) cells were used throughout the study. For the induction of differentiation, PC12 cells were seeded on PDL/Laminin coated glass coverslips or PDL/Laminin coated Si patterned surfaces and left 24 h to adhere and interact with the substrate. The following day 50 ng/ml NGF in DMEM supplemented with 2 % HS was added for the indicated time points. For the phosphorylation analysis of Myosin II Light Chain before the addition of NGF, the cells were serum starved for 4 h, the NGF was added diluted in serum free DMEM and incubated for the indicative time points.
2.5. Immunohistochemistry
For the PC12 cells fixation, a 10X Fixative solution (20 % formaldehyde, 2 % glutaraldehyde, 70.4 mM Na2HPO4, 14.7 mM KH2PO4, 1.37 M NaCl and 26.8 mM KCl) was diluted to 1X with PBS++. Cells were fixed for 10 min with prewarm 1X fixative solution and washed three times with PBS++. For permeabilization, the cells were treated with 0.1 % Tritox-X100 in PBS++ for 5 min. Blocking step was performed with 2 % BSA for 30 min and the primary antibodies were incubated for 1 h at RT or o/n at 4 °C in case of YAP. After 3 washes with PBS++, the secondary antibodies were added for 45 min at RT. After 3 washes with PBS++, the samples were mounted on glass slides with DAPI and observed with Confocal Laser Scanning Microscope (Leica TCS SP8) using a HC PL APO CS2 40x/1.30 NA or 63x/1.40 NA oil lens. The pinhole size was set to 1AU.
2.6. Scanning electron microscopy
PC12 cells were fixed with a mix of 2.5 % GDA and 2.5 % PFA solutions for 15 min and washed 3 times with 0.1 M Sodium Cacodylate buffer. Next, they dehydrated in a series of ethanol solutions (30 %, 50 %, 70 %, 80 %, 90 % and 100 %) for 10 min. Finally, two incubations for 5 min with ethanol:HMDS 2:1 and 1:1 and a last wash with 100 % HMDS were performed and the samples were left o/n to air dry. Before observation, sputter-coating with 15 nm layer of gold was performed and the specimens were observed with SEM (JEOL JSM-6390 LV, Jeol USA Inc, Peabody, MA, USA) with an acceleration voltage of 15 kV.
2.7. Image acquisition and quantitative image analysis
For the quantification of confocal images, once the imaging settings were selected were kept constant during acquisition for all samples. Quantitative image analysis was performed with the ImageJ software (U. S. National Institutes of Health, Bethesda, Maryland, USA, https://imagej.nih.gov/ij/). The pMLC+ cells were manually selected and counted with the Cell counter plugin of ImageJ. Their corrected total cell fluorescence (CTCF) was calculated using the following formula: CTCF = Integrated Density - (Area of selected cell x Mean fluorescence of background readings). The YAP nuclear to cytoplasm intensity ratio was automatically calculated using the Intensity Ratio Nuclei Cytoplasm Tool for ImageJ (Intensity Ratio Nuclei Cytoplasm Tool, RRID:SCR_018573).
2.8. Statistical analysis
All experiments were performed at least three times through the study. At least 100 cells across 3 independent experiments were used for the quantification analysis of YAP and pMLC. For each experiment, one-way ANOVA and Tukey HSD post hoc tests was performed using GraphPad Prism Version 9.0.0 (121) (GraphPad Software, San Diego, California USA, www.graphpad.com). P-values <0.05 were considered statistically different and marked by ∗p < 0.05, ∗∗p < 0.01, or ∗∗∗p < 0.001.
3. Results and discussion
The PC12 cell line is a classical neuronal cell model due to its ability to acquire sympathetic neurons features when treated with NGF. Undifferentiated PC12 cells are small and round, with no neurites and can proliferate indefinitely. Shortly after the addition of NGF, the PC12 cells cease proliferation and start to differentiate. After 3-5 days, most cells are multipolar and bear neurites that extend in multiple directions, reaching other neurites and creating junctions with adjacent neurons. The result is the formation of a complex network with randomly oriented multi-branching neurons.
In our previous work [17], PC12 cells were cultured on laser patterned Si surfaces decorated with periodic arrays of microcones. Three types of micro conical morphologies exhibiting different geometrical characteristics were used along with the flat Si surface. These three surfaces were referred as to low, medium and high roughness. The topological characteristics of these patterned surfaces were described in detail [17]. Briefly, from the analysis of the geometrical characteristics, it seems that as the roughness increases the height of the microcones increases by a factor of two as well as the interspace distance. Hence, the density of the microcones decreases as the roughness increases.
Our previous results showed that when PC12 cells are cultured on Si substrate with high roughness for 4 days, the differentiation ratio is reduced by more than six times, or by more than four times compared to the low and medium roughness respectively. On the 7th day the differentiation on high roughness is completely inhibited. Moreover, the neurite length is reduced three times for the cells that grew on high roughness compared to the length of the neurites of cells that grew on low roughness. As all the different Si topographies were structured on a single substrate, which was placed in a single culture well, as shown in Fig. 1, we safely concluded that the underlying topography is responsible for the alteration in cell behavior [17,29]. Therefore, we sought to investigate this further by focusing on the early differentiation events, i.e., only 24 h after the continuous induction with NGF, which are critical to the progression of the process.

Fig. 1. A: Illustration of Si samples with planar Si and laser micropatterned Si areas decorated with microcones with different geometrical characteristics (low, medium and high roughness). B: SEM images of the surfaces of flat Si and micropatterned Si with low, medium and high roughness. |
3.1. Cell and growth cone morphology analysis
Initially, the morphology of PC12 cells, cultured on the different substrates and continuously treated with NGF for 24 h, was analyzed by SEM (Fig. 2). Low magnification images (Fig. 2A) on glass coverslips (control substrate) (Fig. 2A a,f), flat (Fig. 2A b,g) and low roughened Si (Fig. 2A c,h) showed that PC12 cells are well spread on the substrate surface, while the cell somas show the normal polygonal shape. On medium roughened substrates (Fig. 2A d,i), most of the cells appear to be bipolar, while on high roughened ones (Fig. 2A e,j) less elongated and more bulged cells are observed. On glass, flat and low roughened Si, cell neurites appear to be long, with extensive branching toward multiple directions. However, as the roughness increases, smaller and fewer neurites are formed. Hence, on high roughness most of the cells show a few small or no neurites at all. Higher magnification images (Fig. 2B) showed that on high roughness the cell somas are laying on top of the microcones but as the rest of the cytoplasm slightly sinks in the space between them, small bulges are formed wherever the microcones tips are located underneath (white arrows in Fig. 2B e, f and g). On low roughness, the microcones have lower aspect ratio and this effect is limited, therefore the cell surface morphology appears smoother (Fig. 2B c) like the surface of the cells on glass and planar Si (Fig. 2B a and b). On medium roughness, the aspect ratio is similar to the high roughness but the interspike distance is half, therefore the surface distortion is also limited (Fig. 2B d). From the above observations it is clear that the PC12 cell differentiation process is altered and the cells morphology is remodeled due to the underlying substrate morphology. Neuritogenesis also seems to be heavily affected by the high degree of roughness. Numerous studies have shown that substrate topography can alter cell attachment, cytoskeletal structure and cell shape and that these alterations can affect downstream signaling pathways by altering cell behavior [30,31,32]. Hence, although initially it was believed that the morphology of the cell is determined by its function accumulating evidence demonstrates that the opposite is also true: physical properties of the cellular microenvironment determine cell shape and lineage fate decisions and thus its function [33,34,35]. Neuritogenesis, the process of neurite sprouting from the cell soma, is the first step towards adopting a mature neuronal morphology and relies heavily on growth cone formation and stabilization. Growth cones are very motile expansions at the distal end of all neurites, capable of sensing the environment and guiding the neurite [36,37,38]. The inability of cells cultured on high roughness Si substrate to adopt this characteristic neuronal behavior, motivated us to investigate further the development of the growth cone of NGF-treated PC12 cells. Representative SEM images of the growth cones in PC12 cells cultured for 24 h on the different substrates used are shown in Fig. 3. Top (Fig. 3A a-e) and tilted view (Fig. 3A f-j) images were obtained for all samples tested. On glass and flat Si (Fig. 3A a,b,f and g) we observed the typical morphology of the growth cone with the palm-like structure of the lamellipodium and the finger-like protrusions of the pointed filopodia. The lamellipodium is firmly attached and spread on the substrate and ends up in a rich area of micro protrusions that freely extend and branch into multiple directions. On the low roughness (Fig. 3A c and h), the lamellipodium is wide and spread on top of the microcones with morphology similar to the ones observed on glass and flat Si. The many filopodia that are observed grow unhindered on the substrate and easily bridge the microcones' tips as they extend through the relatively small gaps throughout the discontinuous surface. On the medium roughness (Fig. 3A d and i), the lamellipodium is also spread on top of the microcones tips. The filopodia although are fewer either they are perched on the microcones' tips, or they are falling and wrapping around the microcones' bodies. On the high roughness (Fig. 3A e and j), the lamellipodium is narrow and cannot easily traverse from one microcone tip to the other, hence it grows only in the flat area between the base of the microcones. No filopodia are seen hanging between the microcones’ tips. The very few and short filopodia that are formed, are rather suspended instead of firmly attached, trying unsuccessfully to reach and adhere to the microcones. It is possible that the inability of filopodia to persist on top of the microcones tips long enough for the stabilization of the growth cone is responsible for the narrow shape of the lamellipodium and the decreased neurite sprouting and branching [39]. Similar results were obtained with fibroblasts [40] where the cell spreading, the lamellipodium area and the number of filopodia were reduced as the surface roughness increased. Moreover, it has been shown that the cell spreading of fibroblasts between deformable surfaces, such as flexible polymeric micropillars, is associated with increased cell contractility, actin cytoskeleton reorientation and cell and nucleus shape deformation [41].

Fig. 2. Representative SEM images of PC12 cells treated with NGF for 24 h on glass, flat Si and micropatterned Si substrate with increased roughness (Low, Medium and High). All substrates have been coated with PLL and laminin prior to cell seeding. The upper panel (A) shows low magnification images, while the lower one (B) shows higher magnification ones. |
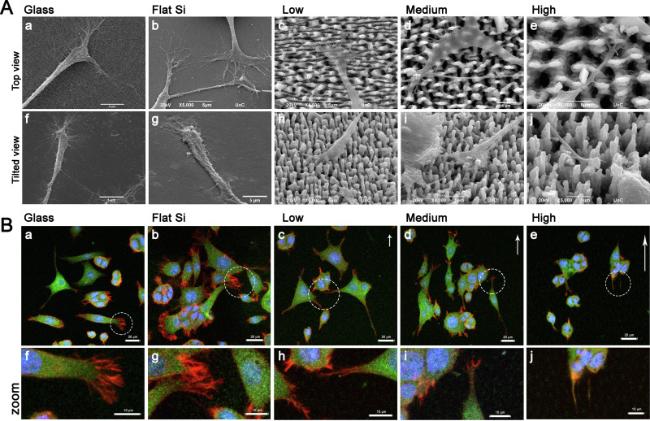
Fig. 3. Analysis of growth cone formation of PC12 cells grown on the different substrates tested. A: Representative SEM images of the growth cones of PC12 cells treated with NGF for 24 h cultured on glass, flat Si and Si substrates decorated with microcones with different roughness (Low, Medium and High) (Scale bar 5 μm). B: Representative confocal microscopy images of PC12 cells treated with NGF for 24 h on glass, flat Si and Si substrates decorated with microcones with different roughness. Cells were stained for β-ΙΙΙ tubulin (green) and F-actin (red). Cell nuclei were stained with DAPI (blue). The white arrows indicate the directionality of the microcones tips; and the length of the arrows represents the degree of the directionality. The areas enclosed by the dashed line in figures a-e in the upper panel, are shown in higher magnification in the lower panel (images f-j). The dark areas present in the protoplasm, resembling holes, are more profound on the cells cultured on the substrate with high roughness, correspond to the microcones tips that lie underneath (See also Fig. 2B). (Scale bar: top panel 20 μm, lower panel 10 μm). All substrates have been coated with PLL and laminin prior to cell seeding. Samples were imaged with a Leica SP8 confocal microscope using a HC PL APO CS2 40x/1.30 NA oil lens. The pinhole size was set to 1AU. |
PC12 cell morphology, neurite outgrowth and growth cone formation were also analyzed by immunocytochemistry and confocal microscopy through the observation of the two major cytoskeletal components, namely tubulin and actin, just 24 h after the addition of NGF. Representative images are shown in Fig. 3B, presenting the overlay images of the triple staining for neuron-specific β-III tubulin (green), F-actin (red) and DAPI (blue). At this early stage of differentiation newly formed neurites still cohabit with lamellipodia making it difficult to discriminate between them. Based on one mechanism of neurite formation there is a collapse of the circumferential lamellipodium in distinct regions while it is extending in others leads to nascent growth cones of what will become newly formed neurites [42,43,44]. However, the results from the confocal microscopy images agree with the information extracted from the SEM images. On glass, flat Si and low roughened Si (Fig. 3B a,b and c), the cells obtained the typical morphology of the differentiated neurons: randomly oriented polygonal and multipolar cells, bearing neurites. Besides, the morphology, the expression of β-III tubulin (green) is additional evidence of their neuronal fate. The typical appearance of the growth cone is also confirmed on these substrates: a growth cone central domain rich in tubulin surrounded by actin-rich lamellipodia and filopodia (lower panel in Fig. 3B f, g and h). However, higher actin expression on the growth cone is observed on the planar surfaces compared to the ones with the micropatterns. On medium roughness (Fig. 3B d and i), most of the cell bodies have obtained an oval shape and the neurites are elongated along the direction dictated by the tips of the microcones. As a result, many cells have become bipolar. The growth cones exhibit normal morphology, although with less dense actin lamellipodia and filopodia. On the contrary, on high roughness (Fig. 3B e and j), although the cells express β-III tubulin most of them remain round in shape or slightly oval following the directionality dictated by microcones’ tips. Only a few or no neurites at all are observed. The neurites, if any, are short and end up in undeveloped growth cones with rudimentary actin lamellipodia. Filopodia are absent or too small to be clearly distinct.
3.2. Analysis of vinculin expression and myosin II ligh chain phosphorylation
As already mentioned, the navigation of the sprouting neurites and the expansion of the growing axons require the growth cone formation and stabilization. Growth cone motility integrates attractive and repulsive molecular cues to navigate the developing axons toward their targets. Among the many cytoskeletal components that are related to the growth cone motility and stabilization, vinculin is important for maintaining strong contact with the substrate and myosin II is responsible for the modulation of the cytoskeletal rearrangement that drives growth cone shape and reorientation. Therefore, next we focused on studying the vinculin expression and the activation of myosin II in PC12 cells cultured on different substrates. The results are depicted in Fig. 4, Fig. 5. In this Fig. 4 we see the typical punctuated pattern of vinculin expression in the cell somas. However, vinculin is also expressed in the growth cones where it is highly recruited in the areas of focal adhesions (FAs, white arrowheads in Fig. 4A and B). It is remarkable to observe that as the roughness of the substrate increases, the total expression of Vinculin but most important its recruitment to the FAs, decreases. Additionally, the size and the shape of the FAs as marked by the vinculin staining, seem to change as the substrate geometry changes. On glass, flat Si and low roughness Si (Fig. 4A a,b and c), the FAs are large and elongated as the clusters of vinculin form the characteristic dashed pattern. This is clearly shown on the higher magnification images in Fig. 4B a and b. On the medium and the high roughness (Fig. 4A d and e), the size of the focal adhesions is smaller and the dot-like FAs are predominant (also in Fig. 4B c and d) These differences in the size and the shape of FAs usually are associated with different maturation stage, with the larger and elongated FAs to be more mature [45].
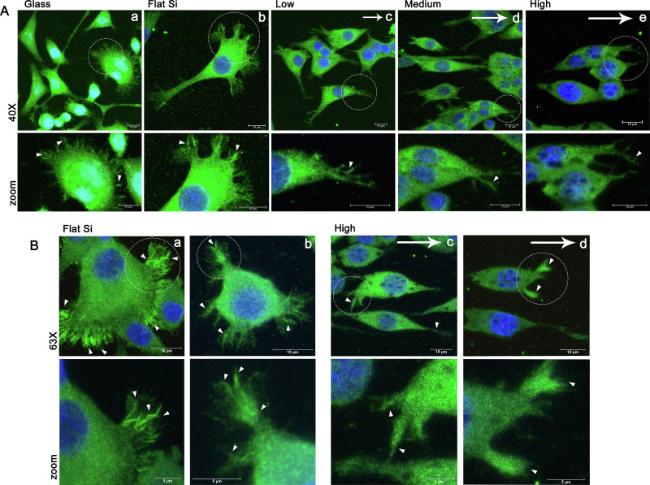
Fig. 4. Vinculin expression in PC12 cells cultured on glass, flat Si and Si substrates decorated with microcones with increased roughness (low, medium and high). A: Confocal microscopy images at 40× magnification of PC12 cells treated for 24 h with NGF and stained for Vinculin (green). Cell nuclei were stained blue (DAPI). B: Confocal images at 63× magnification of PC12 cells cultured on flat and high roughened Si and treated with NGF for 24 h. The lower panel (zoom) shows the selected areas of the images in the upper panel in higher magnification. The arrows indicate the directionality of the microcones of the substrate and the arrowheads indicate focal adhesion points. Samples were imaged with a Leica SP8 confocal microscope using a HC PL APO CS2 40x/1.30 NA and 63X/1.40 oil lens. The pinhole size was set to 1AU. |
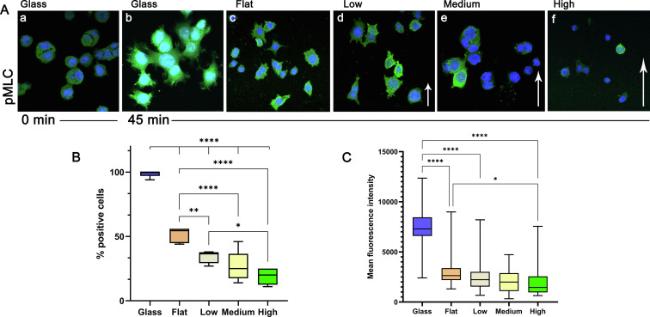
Fig. 5. Myosin II Light Chain phosphorylation (pMLC) in PC12 cells in cultured on glass, flat Si and Si substrates decorated with microcones with increased roughness (low, medium and high). A: Representative confocal microscopy images of PC12 showing the phosphorylation of Myosin II light chain (green) 45 min after the addition of NGF. Cells at 0 min were used as negative control. The dark areas in the protoplasm of the cells correspond to the microcones tips underneath. B: Quantification results of the phosphorylation of Myosin IIB light chain expressed as the % percentage of positive cells. C: Quantification results of the phosphorylation of Myosin IIB light chain expressed as the mean fluorescence intensity of the positive cells. (∗∗p < 0.01, ∗∗∗p < 0.001). Samples were imaged with a Leica SP8 confocal microscope using a HC PL APO CS2 40x/1.30 NA oil lens. The pinhole size was set to 1AU. |
For the neurite sprouting and outgrowth, two components of the growth cone are necessary: the formation of actin-rich filopodia and lamellipodia and the exertion of contractility force on the substratum [46]. Growth cones pull their neurites forward during outgrowth (Retrograde flow, RF), through myosin II-depended contractile mechanisms. Non-muscle myosin II is the most important traction force generator (cell tension) in this process. The observation so far that on higher roughness, fewer and shorter neurites are formed, with narrow lamellipodia and fewer filopodia, raises questions about the activation of myosin IIB inside the cells. Hence, we examined the activation of non-muscle myosin II via phosphorylation of its regulatory light chain (MLC). A pulse-chase experiment of PC12 on glass was conducted and the phosphorylation of MLC was evaluated with confocal microscopy at 0, 5, 15 and 45 min after the addition of NGF. The 45-min time point was selected for the rest of the samples. In Fig. 5A representative confocal images for the detection of the phosphorylated form of MLC (pMLC) in PC12 cells cultured on different substrates and treated with NGF for 45 min are shown. PC12 cells on glass at 0 min are used as the control culture (Fig. 5A a). In Fig. 5B and C the results for the quantification of the phosphorylation of MLC are presented. The percentages of the positive cells regarding the pMLC expression and the mean fluorescence intensity (MFI) of the pMLC positive cells are given in Fig. 5B and C respectively. The MFI of the cells at 0 min was used as the threshold. The results show that on glass, 45 min after the addition of NGF (Fig. 5Ab, graphs 5B and 5C), in almost all the cells the MLC is heavily phosphorylated, hence the high intensity of the staining. On flat Si (Fig. 5A c and 5B), about half the cells (51.6 ± 5.60) have phosphorylated MLC but also the MFI in these cells is almost 3 times lower (Fig. 5C) compared to the MFI of cells on glass. On low roughness only the 33 % ± 5.52 of the cells are pMLC + but the MFI is comparable to the MFI of the cells on flat Si (Fig. 5B and C). As the roughness increases the percentages of the pMLC+ cells decrease to 26.6 % ± 11.92 and to 19 ± 5.89 % for the medium and high roughness respectively (Fig. 5B). The differences in the percentages of the pMLC+ cells on flat Si and on the micro-structured Si are statistically significant (p < 0.001) as well as the differences between the micro-patterned Si with low and high roughness (p < 0.01). Although the differences in the MFI of the cells that are grown on Si are not statistically significant, a trend is observed that shows lower values of MFI as the roughness increases (Fig. 5C). The results from the activation of MLC showed that as the roughness increases the cell tension decreases.
Summarizing the findings so far, we have demonstrated that as the substrate roughness increases the cells have fewer and shorter neurites, the growth cones are undeveloped with narrow lamellipodia and fewer filopodia, vinculin recruitment at focal adhesions is limited and the activation of MLC upon NGF triggering is reduced. In the literature there are many biomaterials and topographical features at the nanoscale that promote the formation of smaller contact points instead of strong focal adhesion points, especially at the growth cone area and this favors neuronal differentiation in different neuronal cell types [47,48,49,50]. On the other hand, in a similar study it was shown that lower number of neurons are formed on surfaces decorated with nanopilars albeit these neurons grew neurites faster these neurites were fewer compared to the flat surfaces [51]. The authors concluded that too distant and small topographical adhesion sites can lower neurite outgrowth. In another study [15] the authors showed that human neuroblastoma cells (SH-SY5Y) when cultured on gold nano-roughened surfaces exhibited reduced adhesion and reduced focal adhesion formation (like the one we see on our Si micro-roughened surfaces) that lead to reduced survival and absence of polarization. In In all these cases, the 3D topographies add o lot of complexity to cell adhesion compared to the flat surfaces making it hard to understand what exactly topographical signal the cell perceives. Hence in similar studies the cell response to topographical cues may seem contradictory, especially when different chemical signals are provided along with the mechanical ones as result of different coating on the culture surfaces.
Growth cone development, focal adhesion formation, stability of actin cytoskeleton, myosin II activation and cell tension are all connected with a key molecule of the substrate-mediated mechanotransduction, coined as the YAP protein. Therefore, we sought to study the expression and the NGF-triggered compartmentalization of YAP in PC12 cells cultured on the different substrates.
3.3. YAP expression and compartmentalization
Initially, a pulse-chase experiment was performed to study the effect of NGF in YAP mobilization (Fig. 6B). PC12 cells were cultured on PDL/laminin coated glass coverslips for 24 h and treated with NGF for various time points (0, 5, 15 and 45 min). Following NGF treatment, the cells were fixed, stained for YAP and observed with confocal microscopy. Representative confocal images are shown in Fig. 6B. The cells without any treatment (Fig. 6B) are round in shape and exhibit a uniform distribution of YAP. Immediately after the addition of NGF, the mobilization of YAP begins, and its nuclear translocation becomes apparent at 15 min. At 45 min a high amount of YAP has accumulated in the nucleus while the cell elongation also starts to become obvious. From this pulse-chase experiment, the time point of 45 min was selected as the most appropriate to calculate the YAP nuclear to cytoplasm intensity ratio for the rest of the samples.

Fig. 6. Expression and NGF-triggered compartmentalization of YAP, 45 min after the addition of the factor in PC12 cells cultured on glass, flat Si and Si substrates decorated with microcones with increased roughness (low, medium and high). A: Representative confocal images of PC12 cultured for 24 h on glass, flat Si and Si substrates decorated with microcones with increased roughness. The cells treated for 45 min with NGF and stained for YAP (green). B: Representative confocal images of a pulse-chase experiment of PC12 cells treated with NGF on PDL/laminin coated glass coverslips. The expression and the nuclear translocation of YAP (green) are shown. C: Quantification of the nucleus to cytoplasm intensity ratio (n/c ratio) of YAP in PC12 cells cultured for 24 h on different substrates and treated with NGF for 45 min (asterisks denote statistical significance: ∗p < 0.05, ∗∗p < 0.01, ∗∗∗p < 0.001). All substrates were coated with PDL/laminin before cell seeding. Samples were imaged with a Leica SP8 confocal microscope using a HC PL APO CS2 40x/1.30 NA oil lens. The pinhole size was set to 1AU. |
PC12 cells were cultured on different substrates for 24 h. The following day, NGF was added for 45 min and then the cells were fixed, stained for YAP and observed with confocal microscopy. Images were taken (Fig. 6A) and the nuclear to cytoplasm intensity ratio (n/c ratio) of YAP was calculated (Fig. 6C) as described in “Materials and Methods” section. From Fig. 6A and C, it is apparent, that in cell cultures on glass, flat Si and low roughness Si substrates, most of YAP has been transported into the nucleus 45 min after NGF addition. The n/c ratio of YAP is 2.72 ± 0.28 for the cells on glass, 2.69 ± 0.55 on flat Si and 2.23 ± 1.12 on the low roughened Si. As the roughness increases the n/c ratio of YAP decreases. For the cells on the medium roughness is 1.33 + 0.42, while on the high roughness is 0.96 + 0.29. This decrease in n/c ratio shows that at higher roughness the cytoplasmic retention of YAP is occurred and hence its nuclear translocation and consequently its activation is hindered.
Taking together, these results show that the substrate geometry, in this case the roughness, influence cell morphology and YAP nuclear localization in the following way: as the roughness increases the cell spreading and the YAP translocation to the nucleus decreases. This is in agreement with other findings where it was shown that the physical restrain of cell spreading leads to retention of YAP to cytoplasm [52]. Many studies so far have confirmed that in 2D cultures increase cell spreading and cell contractility, formation and stabilization of FAs and translocation of YAP/TAZ to the nucleus [52,53,54,55,56] are all connected with stiffer substrates. One possible explanation of this behavior of YAP on stiff substrates is that cell spreading leads to nuclear flattening and stretching of the nuclear pore which enhance YAP/TAZ nuclear import [57]. In addition, it has been showed that rigidity induces YAP phosphorylation and subsequently Smads phosphorylation and therefore their retention in the cytoplasm which in turns results in differentiation of human pluripotent stem cells in motor neurons [58]. Similarly, soft hydrogels inhibit YAP nuclear translocation and promote highly efficient differentiation of hPS cells into postmitotic neurons [59]. These results agree with a study of Baek et al. where cytoplasmic YAP retention coincides with high differentiation of human neural stem cells into neurons cultured on high-resolution nanogroove structure with extremely narrow contact width (15 nm) [50].
However, other biophysical cues such as substrate directionality and architecture that alter the cell shape, could act on YAP signaling apart from stiffness but these remain more undefined. In addition, it has been shown that in more realistic 3D microenvironments YAP/TAZ translocation can increase with stiffness, decrease or remain unchanged thus exhibit counterintuitive trends [52,60,61]. In general, the nuclear translocation of YAP has been correlated with cell survival, proliferation or differentiation towards specific lineages, but this response depends on the cell types and the culture conditions.
In this work, we demonstrated the association of topography in the mechanotransduction process for the alteration of the pattern of NFG-induced differentiation of PC12 cells. Specifically, when PC12 cells cultured in the presence of NGF on Si substrates decorated with microcones with different roughness, the differentiation process is altered and their morphology is remodeled due to the underlying substrate topography. We observed that as the substrate roughness increases, the cells despite expressing neuron-specific βΙΙΙ tubulin, are smaller and rounder in shape, they have fewer and shorter neurites, while the respective growth cones appeared to be undeveloped with narrow lamellipodia and fewer filopodia. In addition, vinculin recruitment at focal adhesions is limited, the activation of myosin II upon NGF triggering is reduced and the YAP nuclear translocation is also limited. SEM analysis showed that the somas of the PC12 cells on highly rough Si, are perched on the microcones tips. Probably, the high discontinuity of the substrate acts as an energy barrier and the cell extension in the horizontal direction is hindered. Therefore, the cells appear smaller, less elongated and more bulged. This barrier has also profound effect on lamellipodium spreading and the formation of filopodia [54,62](Fig. 7). The cells on low roughness can overcome topographical features and the cell somas as well as their protrusions (lamellipodia and filopodia) are well spread on top of microcones. On high roughness, the lamelipodia appear narrow as they cannot traverse from one microcone tip to the other and forced to grow in the defined space near the bases of the microcones. For the same reason, the few filopodia are short and suspended. Due to this situation, the cells on high roughness have fewer contact points with the substrate and less FAs are formed, especially in the growth cone area, which leads to fewer and shorter neurites. The geometry of the substrate affects not only the quantity but also the quality of the FAs. On low roughness, the cells are firmly attached to the substrate due to the increased contact points. This probably creates enough force though the actin-myosin contraction, to the initial adhesion points and expose the vinculin binding sites [63,64]. Hence, vinculin is highly recruited and the adhesion points mature into focal adhesion points. This induce positive feedback and as the tension increases more vinculin is recruited at Fas [64]. As the substrate roughness increases, fewer contact points are established, weaker forces are applied and the vinculin recruitment is limited. These events reduce the retrograde flow (RF) at the growth cones which is responsible for the axonal growth and result to destabilization and retraction of the newly formed axons [65]. This could explain the absence or the short length of the neurites in cells on high roughness. The weaker traction forces inside the cells on high roughness are reflected in the reduced phosphorylation of myosin II upon NGF stimulation and the reduced YAP translocation to the nucleus. All these observations are summarized and depicted in Fig. 7. Our results support the hypothesis that the substrate topography is an important factor affecting cell's morphology and function in manner that under specific circumstances topographical cues can act superiorly to chemical signals. In our case, we cannot rule out the possibility that on the high roughness substrate different opposite effects are on play: the chemical signal from the presence of NGF and the physical barrier from the low density microcones. This opposite effect could be responsible for the mixed phenotypic characteristic of the PC12 cells and therefore we can hypothesize that although NGF induced the differentiation of the cells the topography altered the cell adhesion and subsequently the neurite outgrowth.
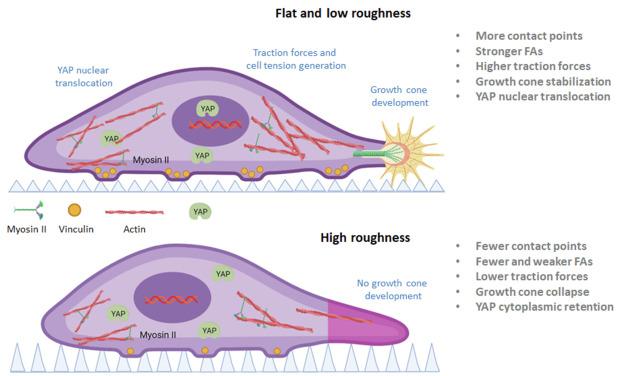
Fig. 7. Illustration of the alterations in mechanosensing and mechanotransduction of PC12 cells cultured with NGF on Si substrates decorated with microcones with different geometrical characteristics. On highly rough substrate, the cell has fewer contact points, especially in the area of the growth cone, and forms less stable and immature focal adhesions. The cellular contractile forces and the axonal retrograde flow are not strong enough to stabilize the growth cone and to promote YAP nuclear translocation. The cell remains round in shape with short or no neurites at all (Created with BioRender.com). |
Others have also shown that microstructure topographical cues can alter PC12 cell differentiation induced by NGF, by modulating neurite outgrowth and guidance, gene regulation, protein expression, etc [28]. Topography-driven signaling can support or impede chemical signaling in an interplay in the level of distinct or intersected and converging intracellular pathways as the cells is exposed to various cues under dynamic environmental conditions [66]. This knowledge is of particular interest in designing topographies to manipulate cell-material interactions toward neural network rewiring in neural tissue engineering applications.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgements
The authors would like to thank Ms. Aleka Manousaki for her valuable help in operating the SEM and acquiring the images. This work was supported by NFFA EUROPE Pilot (EU 2020 framework programme) under grant agreement no.101007417 from 1/03/2021 to 28/02/2026.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}