
1. Introduction
As a significant public health problem with high morbidity and mortality worldwide, cancer is a primary disease that endangers human life [1,2,3]. The dynamic changes of the solid tumor microenvironment (TME) formed by the interaction of tumor cells with their microenvironment are closely related to tumor progression. The TME not only contains tumor cells, endothelial cells, fibroblasts, and various immune cells but also includes the formation of tiny blood vessels and various cell-secreted factors, and the evolution of these components provides a "seedbed" [4]. Early studies focused on the effects of various cell types in TME on tumors and how to target tumor cells in TME to cure them. However, many studies have shown that most solid tumors are accompanied by altered mechanical factors during growth and play an essential role in tumor development. The current mechanical microenvironments that influence tumor progression are (1) solid stress, (2) fluid pressure, (3) matrix stiffness, and (4) physical microstructure [5,6,7]. The extracellular matrix (ECM) is a secretory network of collagen, fibronectin, laminin, and other components that provide structural support and mechanical signals for cell growth [8,9]. In most solid tumors (e.g., hepatocellular carcinoma, breast cancer, and colorectal cancer), increased ECM deposition and collagen crosslinking increases matrix stiffness [10]. The increased matrix stiffness plays a positive role in the biological functions of tumor cells, including cell proliferation, invasion, metastasis, angiogenesis, metabolic reprogramming, and immune escape [11,12].
With the widespread use of elastography and atomic force microscopy (AFM), matrix stiffness is being studied in depth in various diseases [13]. The AFM is the primary instrument for measuring matrix stiffness and is commonly used in biology, medicine, chemistry, and materials science, providing solutions for measuring micromechanics (e.g., cell mechanics) [14]. In contrast, the clinical use of ultrasound or Magnetic resonance imaging (MRI) elastography is often used as an effective adjunct in diagnosing various diseases such as liver fibrosis and solid tumors [13,14,15,16,17,18]. Differences in measurement techniques make the specific stiffness values vary considerably in multiple conditions. However, studies have shown that the stiffness of solid tumor tissue is significantly higher than that of the corresponding normal tissue (Table 1) [19,20,21,22,23,24,25,26,27,28,29]. The stiffness of normal liver tissue is usually less than 6 kPa, while the stiffness of liver cancer tissue exceeds 8 kPa and can even reach over 70 kPa [19,20]. Breast cancer tissue is ten times stiffer than normal breast tissue [21,22]. The advanced stages of hepatitis eventually lead to cirrhosis and liver cancer and are known as the trilogy of liver lesions. Interestingly, Sun et al. utilized chemicals to trigger hepatitis, cirrhosis, and liver cancer in rats. They then measured the stiffness of the liver tissues using AFM and observed a progressive increase in tissue stiffness as the diseases advanced [30]. In bladder cancer, the matrix stiffness of recurrent tumor tissue was significantly higher than that of tumor tissue from new patients [24]. This was also verified in gliomas, where matrix stiffness was significantly higher in highly malignant gliomas (10 kPa) than in low-grade malignant (1 kPa) and non-malignant glioma tissue (0.1 kPa) [31,32]. These studies suggest that matrix stiffness is essential in promoting tumor progression.
Table 1. The stiffness of human normal tissues and the cancerous tissues. |
| Tissues | Types | Stiffness (kPa) | Refs. |
|---|---|---|---|
| Liver | Normal | <6 | [19,20] |
| Tumor | >8, up to 70 | ||
| Breast | Normal | about 1.13 | [21,22] |
| Tumor | up to 55 | ||
| Thyroid | Benign | about 20 | [23] |
| Malignant | ≥28.2 | ||
| Colorectum | Normal | <5 | [26] |
| Tumor | about 7.51 | ||
| Bladder | Normal | <5 | [24] |
| Tumor | >5(Up to 10) | ||
| Endometrium | Normal | about 23.8 | [25] |
| Tumor | about 39.4 | ||
| Skin | Normal | 0.401 ± 0.148 | [27] |
| Tumor | 0.787 ± 0.056 | ||
| Prostate | Hyperplasia | 56.7 ± 14.0 | [28] |
| Tumor | 92.8 ± 21.5 | ||
| Pancreas | Normal | 5.66 (4.39-7.84) | [29] |
| Tumor | 5.70 (3.48-11.55) |
The combined action of tumor and stromal cells, which secrete large amounts of collagen and stromal cross-linkers, is the main reason for the increased matrix stiffness in the tumor microenvironment [33,34,35,36]. The increased matrix stiffness provides a comfortable environment for tumor growth and increases tumor cell proliferation, migration, and invasion [37]. In addition, tumor cell growth leads to TME changes that activate cancer-associated fibroblasts (CAFs) to secrete collagen and fibronectin, further increasing matrix stiffness in tumor tissue [14,38,39]. It is suggested that a positive feedback loop may exist to promote tumor progression. According to existing studies, matrix stiffness can modulate multiple signaling axes to promote tumor development. However, the molecular mechanisms of how matrix stiffness regulates tumor progression and how tumor cells sense and respond to mechanical signals are unclear. Recently, Hanahan et al. has recapitulated the characteristics of the tumor, allowing us to have a more straightforward and deeper understanding of the characteristics of the cancer [40,41]. Numerous studies have shown that matrix stiffness is critical to these abnormal tumor processes (Fig. 1). In this review, we summarize the detailed mechanisms by which matrix stiffness promotes tumor progression and the therapeutic modalities that may alter matrix stiffness, with the expectation that targeting matrix stiffness will provide new approaches to treat tumors.

Fig. 1. Matrix stiffness regulates the function of cancer cells. Matrix stiffness plays an active role in tumor cell biological function. Tumor cells benefit from elevated matrix stiffness to promote autogenous growth, migration, and evasion of treatment. |
2. Hallmark of matrix stiffness-driven cancer progression
2.1. Matrix stiffness maintains tumor proliferation-related signals
Cell proliferation is required during normal tissue development and maintenance and is regulated by sophisticated regulation of multiple signals inside and outside the cell. However, uncontrolled regulation of these signals in tumor cells leads to rapid cell proliferation. It has been shown that matrix stiffness promotes tumor cell proliferation by altering multiple intra- and extracellular signals.
Nuclear factor-κB (NF-κB) is a crucial regulator of tumor development and is essential in tumor initiation, proliferation, and progression [42,43,44]. In lung cancer cells, higher matrix stiffness promotes tumor cell proliferation through actin contraction induced by phosphorylation of myosin regulatory right chain (MRLC), thus activating NF-κB [45]. Activation of the serine/threonine kinase/mammalian target of rapamycin (AKT/mTOR) signaling pathway can strongly induce the proliferation of tumor cells. Higher matrix stiffness can increase AKT activity through the oncogene ZNF217, activating the Phosphatidylinositide 3-kinases/Rac (PI3K/Rac) and extracellular regulated protein kinases (ERK) pathways and promoting tumor cell proliferation [46,47]. In addition, CXC chemokine receptor (CXCR), a member of the G protein-coupled receptor superfamily, is composed of CXCR1-7 small molecule proteins, and plays an essential role in tumor proliferation by binding to its respective specific ligands [48]. Matrix stiffness can regulate the growth of hepatocellular carcinoma (HCC) cells by regulating CXCR4 expression and through the YAP signaling pathway, further suggesting that increased matrix stiffness can promote proliferative signaling in HCC [49]. Activation of the Yes-associated protein/PDZ-binding motif (YAP/TAZ) signaling pathway can promote tumor cell proliferation by regulating the cell cycle, DNA replication and repair, mitosis, and other factors [50]. It was shown that the proliferation rate of soft nest carcinoma cells SKOV-3 cultivated on a stiff matrix was significantly faster than on a soft matrix. In addition, the expression of YAP was markedly higher in cells under higher matrix stiffness, suggesting that the YAP signaling pathway plays an active role in the proliferation of tumors under higher stiffness [51]. Moreover, several studies of hepatocellular carcinoma found that increased matrix stiffness could activate the YAP/TAZ signaling pathway, promote the proliferation of hepatocellular carcinoma cells, and thus promote the progression of hepatocellular carcinoma [49,52,53,54].
Furthermore, tumor cell proliferation is influenced by matrix stiffness regulating non-tumor cells in the tumor microenvironment. Under higher matrix stiffness, MSCs can differentiate into CAFs and secrete the soluble factor prosaposin, promoting breast cancer cell proliferation [55]. Higher matrix stiffness also induces autophagy in fibroblasts and stellate cells, promoting the growth of adjacent tumor cells [56].
Thus, higher matrix stiffness can promote proliferation by modulating the tumor cell or microenvironment. Interestingly, matrix stiffness can also promote the proliferation of non-tumor cells, such as vascular endothelial cells [57], Mesenchymal stem cells [58,59], and airway smooth muscle cells [60]. The growth of multiple cells was promoted at high stiffness, suggesting that matrix stiffness is a potential target for treating proliferative diseases.
2.2. Matrix stiffness promotes tumor invasion and metastasis
Tumor invasion and metastasis are the leading cause of death in tumor patients. Studies have shown that the metastatic tumor tissue matrix was significantly stiffer than the primary tumor matrix [31,32]. The change in matrix stiffness can be sensed by tumor cells by the mechanical sensor, and tumor cells respond with a series of pathological changes promoting their own motility and invasive ability [61].
YAP/TAZ is the most studied transcription factor regulated by matrix stiffness and is associated with the invasion and metastasis of many tumors. Matrix stiffness can regulate YAP/TAZ expression through multiple molecular mechanisms. Higher matrix stiffness regulates Rho-kinase (ROCK) signaling, leading to phosphorylation of MRLC and contraction of myosin, which translocate YAP/TAZ into the nucleus of tumor cells, thereby activating YAP/TAZ to promote tumor invasion and metastasis [62,63,64]. Interestingly, there is a positive feedback loop between ROCK signaling and YAP activation in tumor cells, with activated YAP promoting ROCK expression. Matrix stiffness can promote tumor invasion and metastasis by promoting CXCR4 expression, thereby reducing the expression of ubiquitin domain-containing protein 1 (UBTD1), capable of degrading YAP, resulting in elevated YAP activity [49]. Matrix stiffness also activates the mitogen-activated protein kinase (MAPK) signaling pathway, which activates YAP into the nucleus of tumor cells, promoting tumor invasion and metastasis [52].
In addition, matrix stiffness regulates β-catenin expression to increase tumor invasion and metastasis. In HCC, a higher matrix stiffness accelerates tumor progression through the integrin β1/GSK-3β/β-catenin signaling pathway. It induces cellular communication network factor 1 (CCN1) expression, promoting β-catenin activity and N-cadherin expression on the endothelial cell surface, enabling tumor cell metastasis [65,66]. Tumor suppressors act as supervisors of multiple cellular processes that inhibit tumor development, but tumors can overcome these barriers through various pathways. Studies have shown that matrix stiffness can inhibit tumor suppressor activity. RASSF1A is a tumor suppressor in non-small cell lung cancer, and its epigenetic silencing is associated with advanced disease, metastatic potential, and poor prognosis [36]. It was shown that the expression of RASSF1A was significantly reduced in lung cancer cells under higher matrix stiffness culture, driving the metastatic progression of lung adenocarcinoma through P4HA2 [36].
Epithelial-mesenchymal transition (EMT) is one of the biological processes of invasion and metastasis of tumor cells. During EMT, tumor cells gradually lose epithelial properties such as cell junctions and polarity and acquire mesenchymal characteristics and invasive capacity. Matrix stiffness can regulate EMT through molecular signals to promote tumor invasion and metastasis. TWIST1 is a highly conserved transcription factor containing a base-helix-loop-helix structural domain, and studies have shown that TWIST1 plays an essential role in tumor metastasis [67]. A significant positive correlation was found between TWIST1 expression and matrix stiffness in tissues from patients with breast cancer [68]. It was shown that matrix stiffness regulates the expression of TWIST1. In breast cancer cells, higher matrix stiffness drives the release of TWIST1 from its cytoplasmic binding molecule G3BP2, which localizes in the nucleus, thereby causing EMT and promoting tumor invasion and metastasis [69,70]. However, after the knockdown of TWIST1, the EMT phenotype was suppressed under higher matrix stiffness, suggesting that matrix stiffness-induced EMT is TWIST1-dependent [70]. Recent studies found that SiHa cells cultured on higher matrix stiffness had a significantly enhanced EMT capacity, and the expression of TWIST1 and miR-106b increased with increasing matrix stiffness.
Nevertheless, the expression of integrin β1 endocytosed DAB2 was the opposite, suggesting that matrix stiffness targets DAB2 and thus regulates EMT by regulating miR-106b expression [71]. Transient receptor potential vanilloid 4 (TRPV4) is a mechanosensitive ion channel. TRPV4 can promote matrix stiffness-induced EMT by enhancing Akt activation and YAP/TAZ translocation into the nucleus [72,73].
Matrix stiffness also regulates the activity and expression of multiple transcription factors, promoting EMT and invasive metastasis. Higher matrix stiffness encourages the expression of cytokine Snail, triggering EMT and metastasis in HCC cells [11]. In lung cancer cells, transcription factor c-Myb and lymphatic enhancing binding factor 1 (LEF1) is activated with increased matrix stiffness, thereby promoting the expression of discoidin domain receptor tyrosine kinase 2 (DDR2), which in turn causes the onset of invasion [74].
2.3. Matrix stiffness promotes tumor-associated angiogenesis
Angiogenesis is tightly regulated in vivo. However, in tumor development, rapidly growing tumor cells promote continuous angiogenesis, providing a rich supply of nutrients and oxygen for tumor progression and the metabolism of waste products and excretion of carbon dioxide [40]. Abundant studies have shown that matrix stiffness is a critical regulator of angiogenesis.
Vascular endothelial growth factor (VEGF) is a critical secretory factor in vascular neogenesis, and studies have shown that VEGF expression is significantly elevated in HCC cells and vascular endothelial cells cultured under higher matrix stiffness [75,76]. Hypoxia-inducible factor 1α (HIF1α) can promote the expression of VEGF in hypoxic tissues and induce vascular neogenesis [77]. It was shown that higher matrix stiffness fosters the expression of HIF1α in glioma cells [31]. In breast cancer, the expression of HIF1α and matrix stiffness were positively correlated [68]. Higher matrix stiffness caused activation of Piezo1 inhibited ubiquitination of HIF1α and promoted HCC angiogenesis [78]. In addition, matrix stiffness also regulates VEGF expression through other pathways. Higher matrix stiffness drives vascular survival in HCC by regulating the integrin αVβ5/Akt/Sp1 signaling pathway, which upregulates VEGFR2 in vascular endothelial cells and VEGF expression in HCC cells [79]. Higher matrix stiffness alters the biological behavior of tumor cells, including proliferation, migration, and cell adhesion, which can promote tumor vascularization [80]. Matrix metalloproteinases (MMP) regulate tumor invasion and metastasis, enabling biological functions such as angiogenesis [81]. Through in vivo and in vitro experiments, Francois et al. found that higher matrix stiffness enhanced MMP activity and promoted vascular neogenesis [80]. By co-culturing lung cancer cells and human umbilical vein endothelial cells (HUVEC) on the matrix at different stiffness levels, Dan et al. showed that higher matrix stiffness promoted the expression of MMPs and angiogenesis-related factors and enhanced angiogenesis [76]. In conclusion, matrix stiffness enhances angiogenesis in various tumors by promoting the expression of vascular endothelial growth factors.
2.4. Matrix stiffness promotes tumor treatment resistance
Decreased sensitivity and resistance to radiochemical therapy are leading causes of oncologic treatment difficulties and oncologic death. Understanding the detailed stroma of oncologic therapy improves patient quality of life and enhances patient survival. Studies have shown that matrix stiffness is a favorable barrier and tool for antitreatment in various tumors. Several animal studies have shown that higher matrix stiffness interferes with the penetration of chemotherapeutic agents, which induces tumor insensitivity to chemotherapy. Chemotherapeutic agents such as adriamycin, cisplatin, and sorafenib are currently the classical oncologic agents. Triple-negative breast cancer MDA-MB-231 cells cultured at higher matrix stiffness inhibited integrin-linked kinase (ILK), decreasing sensitivity to adriamycin [82]. In ovarian cancer cells, higher matrix stiffness increased the expression of Focal adhesion kinase (FAK), Akt, and signal transducer and activator of transcription 3(STAT3) in tumor cells, which promoted the activation of YAP, leading to a significant decrease in drug sensitivity to cisplatin [83]. Interestingly, consistent results were obtained in hepatocellular carcinoma [84]. Sorafenib is currently the first-line drug for hepatocellular carcinoma, and the higher matrix stiffness can activate the integrin-Jun N-terminal kinase(JNK) signaling pathway in hepatocellular carcinoma cells, which in turn can develop resistance to sorafenib [85,86]. Metformin is traditionally used in diabetes, but recent studies have shown its therapeutic effects in various tumors [87]. It was found that matrix stiffness reduced PTEN expression and AKT activity, reducing sensitivity to metformin [88]. Radiotherapy as adjuvant therapy is now commonly used to induce tumor cell death and slow tumor growth by stimulating the production of free radicals and reactive oxygen species, disrupting the DNA double helix [89]. The current study found that MDA-MB-231 cells cultured at higher matrix stiffness developed resistance to high radiotherapy [90].
Cancer stem cells (CSCs) are a subpopulation of cells in tumors that are in a stem cell state and have the unique ability to self-renew and differentiate [91,92]. It is one of the leading causes of drug resistance and tumor recurrence [93,94]. SOX2 gene plays an essential role in the self-renewal of CSCs. After laryngeal squamous carcinoma, Hep2 cells were cultured under soft matrix stiffness (1 kPa) or higher stiffness (8 kPa). The expression of SOX2 was significantly higher in cells cultured under soft matrix stiffness, indicating that stemness was maintained in laryngeal squamous carcinoma cells under soft stiffness [95]. Consistent results were also obtained in osteosarcoma cells, with increased expression of the cellular stem cell markers SOX2, OCT4, and NANOG at soft matrix stiffness. They significantly reduced sensitivity to the drug doxorubicin [96]. However, in hepatocellular carcinoma HCCLM3 and Huh7 cells, the investigators found that higher matrix stiffness promoted the expression of the stem cell phenotypes NANOG, OCT4, and CD133 [97]. Consistently, the sensitivity of hepatocellular carcinoma cells to sorafenib was significantly reduced at higher stiffness compared to soft matrix stiffness [97]. It implies that tumor cells' self-renewal ability and drug resistance were enhanced considerably at high matrix stiffness, and their stemness capacity was significantly enhanced. Interestingly, the study by Li et al. found that with increasing matrix stiffness, the proliferation capacity of hepatocellular carcinoma stem cells was enhanced considerably, drug sensitivity was significantly reduced, and the expression of stem cell phenotype markers CD44, CD133, EpCAM, c-Myc, and OCT4 was increased considerably [54]. Sun et al. measured Young's modulus in the invasion front region and core region of hepatocellular carcinoma tissue by atomic force microscopy and combined with immunofluorescence to stain markers of hepatocellular carcinoma stem cells and found that the number of CSCs was also significantly increased in regions with higher Young's modulus [30]. These results suggest that the variation of matrix stiffness is beneficial to the maintenance of stemness characteristics of tumor cells and CSCs. The effects of stiffness on stemness characteristics in different tumors under different matrix stiffness may be related to two-dimensional matrix stiffness and three-dimensional matrix stiffness culture. However, the differences and molecular mechanisms still need further study and exploration.
Even though numerous studies have shown that higher matrix stiffness is resistant to chemoradiotherapy, a few studies have obtained the opposite result. It has been found that higher matrix stiffness enhances the activity of the pro-apoptotic JNK pathway and thus makes breast cancer cells sensitive to chemotherapy. In comparison, soft matrix stiffness gives the opposite result, making tumor cells resistant to chemotherapy [98]. In addition, Lacombe et al. found no significant difference in the sensitivity of melanoma cells SK-MEL-28 and prostate cancer cells PC3 to radiotherapy radiation with altered matrix stiffness, but this may be due to the difference between the plant scaffold culture model and the commonly used hydrogel model [99]. Overall, matrix stiffness-regulated resistance to chemoradiotherapy is currently controversial and may be related to the type of cells and culture mode.
2.5. Matrix stiffness regulates the metabolic reprogramming of tumors
The mode of glucose metabolism in mammalian cells is aerobic phosphorylation. However, the predominant metabolic method in tumor cells under aerobic and anaerobic conditions is glycolysis, referred to as the Warburg effect [103]. The Warburg effect favors tumor progression by rapidly providing nutrients to the tumor. In addition, it is associated with multiple aberrant signals, including Phosphoinositide 3-kinase (PI3K)/AKT signaling pathway, Wnt/β-catenin, HIF1α, etc., thus regulating the expression and activity of metabolism-related proteins [42]. Matrix stiffness is a crucial regulator of tumor metabolic reprogramming, which can affect glucose metabolism by increasing the expression of key glycolytic enzymes through direct or indirect effects. Studies have shown that higher matrix stiffness activates the MAPK-YAP signaling pathway, increases the expression of glycolytic enzymes HK2 and LDHA, and promotes glycolysis in hepatocellular carcinoma cells, leading to tumor progression [52].
Meanwhile, higher matrix stiffness promotes the expression of Glut1 and MCT4 in tumor-associated matrix cells, increases lactate production and glucose uptake, and stimulates the migration of epithelial cells [104]. In addition, matrix stiffness can directly upregulate many metabolism-related factors, such as β-catenin [65,66], HIF1α [31,68], and c-Myc [54]. By regulating these factors, matrix stiffness may affect metabolism in tumors, but more experiments are still needed to verify the relevant findings.
In addition to glucose metabolism, tumor metabolic reprogramming also involves changes in many other critical metabolic pathways, including lipid metabolism, amino acid metabolism, nucleotide metabolism, etc. [105,106,107,108]. Higher matrix stiffness promotes the expression of stearoyl CoA desaturase 1 (SCD1) in HCC cells, altering cellular lipid composition and enhancing HCC cell invasion and metastasis [109]. In tumors, the demand for proline was significantly increased, and in addition, pyrroline-5-carboxylate reductase 1 (PYCR1), a key enzyme for proline synthesis, was significantly overexpressed in tumors [110,111]. Higher matrix stiffness promotes the translocation of kindlin-2 to mitochondria. Translocated kindlin-2 interacts with PYCR1 and promotes PYCR1 expression, leading to increased proline synthesis and tumor cell proliferation [111]. These studies suggest that targeting matrix stiffness may effectively inhibit tumor progression by modulating metabolic reprogramming.
2.6. Matrix stiffness promotes the immune escape of cancer cells
The immune system can prevent tumorigenesis and progression, and evasion of immune surveillance is essential for tumor overgrowth. Several studies have shown that matrix stiffness regulates the immune response.
Programmed cell death ligand 1 (PD-L1) plays an essential role in tumor cell evasion from immune system surveillance. Higher matrix stiffness was shown to promote PD-L1 expression in breast and lung cancer cells [112,113]. Matrix stiffness can affect the distribution of YAP/TAZ, and TAZ can enhance PD-L1 expression by promoting the binding of transcription factors and PD-L1 [114,115].
T-cell activation and expansion are vital drivers of immunity in tumors. In contrast, progressively rising matrix stiffness in tumor development inhibits T-cell activation and infiltration, leading to the immune escape of tumor cells [116]. It was found that the proliferative activity of T cells was significantly reduced at higher matrix stiffness [117]. In addition, the ability of T cells to kill tumor cells was significantly reduced at higher matrix stiffness [118]. Interestingly, in a pancreatic ductal carcinoma model, the investigators found that reduced matrix stiffness greatly enhanced T cells' migration and infiltration rate. The number of CD8+ T cells also increased significantly [119]. Matrix stiffness also regulates the activity of other immune cells. Dendritic cells are critical for T cell activation and immune response induction, and studies have shown that increased matrix stiffness inhibits the ability of dendritic cells to activate immune responses in vitro and inhibits dendritic cell migration, leading to failure of antitumor immunity [120,121,122]. NK cells can kill tumor cells, thus releasing antigens. At the tumor initiation stage, NK cells migrate to the tumor site and release different protein hydrolases to kill tumor cells. However, in higher matrix stiffness, NK cell migration is blocked from reaching the tumor site [123]. Tumor-associated macrophages (TAMs) are the primary cells of the tumor microenvironment. They are often converted to the M1 type in the tumor initiation stage, which can phagocytose tumor cells and inhibit tumor progression. While in tumor progression, various inflammatory factors can promote the conversion of TAMs to M2 type, which can hinder the function of T cells and promote tumor progression [124,125]. Immune components of TAMs are currently shown to be influenced by matrix stiffness [126]. Taufalele et al. found that the higher matrix stiffness region contained a higher proportion of macrophages, mainly of the M2 type, by single-cell analysis of cell subsets in different matrix stiffness regions of breast cancer tissues in the MMTV-PyMT mouse model. The results were validated by in vitro experiments, in which mammary tumor cells cultured under higher matrix stiffness could secrete CSF-1, a protein that promotes macrophage differentiation to M2 [127]. In vivo, experiments also revealed increased M2-type polarization of macrophages in tumor areas under higher matrix stiffness. In addition, these mice with higher matrix stiffness had significantly increased tumor size and were prone to metastasis [128].
Overall, matrix stiffness inhibits immune cells' function and leads to tumor cells' immune escape, which results in ineffective immunotherapy. Therefore, targeting matrix stiffness and degrading collagen fibers may be a future strategy for adjuvant immunotherapy.
3. Targeting matrix stiffness is a potential treatment strategy for solid tumors
The treatment of tumors remains a current challenge in the world of medicine. A variety of current treatment options are ineffective for recurrent or metastatic tumors. There is an urgent need for a novel tumor treatment modality to improve the rate of tumor treatment. Matrix stiffness plays an essential role in tumor progression. Gradually rising matrix stiffness prevents the penetration of anti-cancer drugs and impairs the function of immune cells, so targeting matrix stiffness may be an effective way to treat tumors (Fig. 2).
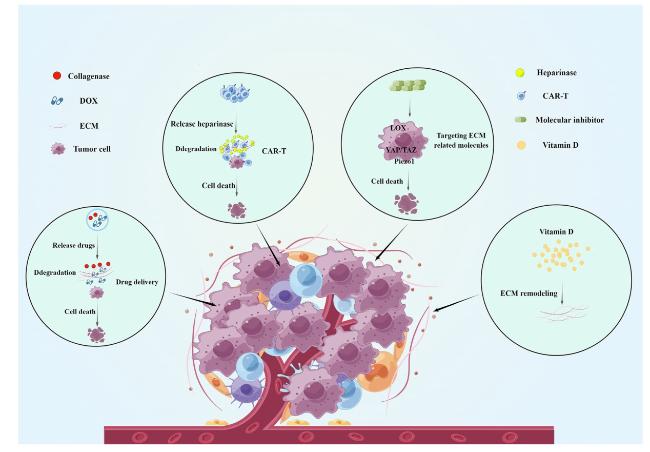
Fig. 2. Targeted matrix stiffness therapy for tumors (By Figdraw). There are currently some practical methods for targeting matrix stiffness, such as drug delivery, targeting matrix-related molecules, etc. These methods have shown promising results in cell and animal experiments, but further clinical research is still needed to verify the therapeutic effect. |
In solid tumors, collagen deposition and crosslinking are leading causes of altered stiffness in the microenvironment. Shou et al. constructed a magnetically ionized modulated hydrogel system that allows dynamic modulation of matrix stiffness, and increased matrix stiffness promotes tumor malignancy, yet softening the matrix at this point reverses tumor progression and promotes the therapeutic effect of doxorubicin [129]. Therefore degrading or altering the deposited collagen may be an effective way to treat tumors. Several studies found that degradation of the extracellular matrix can facilitate drug penetration and thus inhibit tumor progression. Liu et al. used nanoparticles encapsulated with collagenase and chloramphenicol. By targeting the tumor location with the acid response, collagenase was released from the nanoparticles in the tumor. It degraded the extracellular matrix, making it easier for chloramphenicol to reach the tumor cells, and the ability of chloramphenicol to kill tumor cells was significantly enhanced [130]. Similarly, Wang et al. used acetylated dextran (AceDEX) as a carrier material for collagenase to target tumors by pH sensitivity. After loading the chemotherapeutic drug, adriamycin can degrade the extracellular matrix, facilitating the penetration and enrichment of antitumor medicines for tumor treatment [131]. Interestingly, in rats with liver fibrosis (one of the main risk factors for hepatocarcinogenesis), degradation of the extracellular matrix favored the amelioration of liver fibrosis by drugs [132].
Chimeric antigen receptor T (CAR-T) cell therapy is a hot topic in tumor immunotherapy. T cells are isolated and purified from the blood of tumor patients, and chimeric antigen receptor vectors that can recognize and activate T cells are transferred into T cells through biomedical engineering technology, significantly affecting the treatment of hematologic tumors. However, CAR-T is ineffective in solid tumors, possibly due to solid tumors' matrix stiffness that hinders CAR-T's penetration. Constructing CAR-T cells capable of reducing matrix stiffness in TME may effectively treat solid tumors. CAR-T cells with high expression of heparinase can degrade ECM, thereby promoting T cell infiltration and inhibiting the growth of tumor cells [133]. Lysovirus therapy has emerged as a novel immunogenic therapy to enhance antitumor immune responses, and Zhang et al. designed recombinant CAR-T cells carrying core proteoglycans and targeting carbonic anhydrase IX (CAIX) to alter the composition of the ECM and reduce tumor burden [134].
In addition, alterations in matrix stiffness are regulated by various molecules, and the development of drugs targeting these regulatory molecules to block mechanotransduction may be a new strategy for treating tumors. Lysine oxidase (LOX) secreted by matrix cells is the main catalytic enzyme for the crosslinking of collagen and elastin, which, together with transglutaminase, promotes the crosslinking of collagen, resulting in an increase in matrix stiffness [135,136,137,138]. Therefore, inhibition of LOX may be an effective way to reduce matrix stiffness. β-aminopropionitrile (BAPN) is a pan-LOX inhibitor, and several studies have shown that BAPN can inhibit the biological processes of tumor cells [139,140]. CCT365623 is also a LOX inhibitor that regulates the natural function of LOX in tumor cells and has also demonstrated significant antitumor activity in animal studies with better efficacy, oral utilization, and tolerability than BAPN [141]. YAP/TAZ is one of the most essential "force-stimulated sensors" mediating the tumor-promoting effects of stromal stiffness, and Verteporfin is the most common YAP inhibitor, with several preclinical results showing that Verteporfin inhibits the development of a variety of tumors [142,143]. Dasatinib is a tyrosine kinase inhibitor widely used clinically for treating leukemia and solid tumors [144,145,146]. It was found that dasatinib could directly inhibit the YAP/TAZ signaling pathway [147,148]. Piezo1 is another mechanical stimulator that plays an essential role in the biological behavior of altered matrix stiffness. GsMTx4, a peptide derived from spider venom, has a selective inhibitory effect on cation permeable channels of the Piezo family, which can inhibit immune escape in tumors, but research on its inhibitory role in tumor development still needs to be explored [149,150].
Interestingly, vitamin D may have the ability to reduce stromal stiffness. Vitamin D is negatively correlated with the prognosis of many tumors, and inhibits tumor growth [151,152,153,154]. These suggest that vitamin D has tumor-suppressive effects. It was found that vitamin D can inhibit the activation of the WNT4/β-linked protein/mTOR pathway, reducing the production of extracellular matrix [155]. In addition, the combined use of vitamin D receptor modulators and gemcitabine also inhibited ECM remodeling [156]. These results reveal that vitamin D may be therapeutic by regulating tumor matrix stiffness, but more in-depth mechanistic studies are still needed. Certain clinical drugs may be connected to the control of matrix stiffness. Sorafenib is currently the standard treatment for HCC. Recent studies have found that Sorafenib can reduce collagen deposition and thus alleviate liver fibrosis [157]. Simvastatin is a common drug for the treatment of cardiovascular diseases. Studies have found that the targeted delivery of simvastatin by sinusoidal endothelial cells can reduce collagen deposition and inhibit the progress of HCC [158]. These classic drugs may achieve therapeutic effects by reducing the stiffness of the tumor matrix, thus developing new applications. In addition, the combination of these drugs may bring more favorable therapeutic effects. However, more animal and clinical experiments are still needed for validation.
These studies suggest that targeting the extracellular matrix and related molecules is beneficial for treating solid tumors.
4. Conclusions and future prospective
Tumor research continues to evolve, and our understanding of the mechanisms of tumor development continues to improve, but the treatment of tumors remains a significant challenge. Traditional therapeutic approaches usually target tumor cells directly. However, studies have shown that the composition and biochemical properties of the tumor extracellular matrix also play an essential role in tumor development, and it is not easy to improve these aspects with existing therapeutic regimens.
Matrix stiffness in the tumor microenvironment promoting tumor development has received considerable attention. Matrix stiffness is significantly increased in a variety of tumors. It affects the function of tumor cells and interferes with radiotherapy's efficacy in tumor treatment. Although matrix stiffness is being studied in depth, the molecular mechanisms by which matrix stiffness regulates tumors are complex, and many details remain unclear (Fig. 3). YAP, ROCK, and Piezo1, as currently recognized mechanical response factors, play essential roles in tumor progression. As described above, YAP plays a vital role in tumor proliferation, invasion, metastasis, angiogenesis, and other biological behaviors [159,160,161]. Various signaling pathways, such as Wnt and MAPK, also regulate YAP [52,160,162]. ROCK and Piezo1 also play essential roles in the biological behavior of tumor progression [138,163,164,165]. These critical molecules and signaling pathways are interconnected and work together to construct a complex and precise molecular network that mechanically regulates the occurrence and development of tumors. More research is needed to understand the regulatory networks involved. However, how matrix stiffness affects tumorigenesis, progression, and efficacy is still a challenge to be solved. In addition, most current studies on matrix stiffness are in vitro experiments, which are two-dimensional systems and do not reflect the organism's structure. In the future, a more mature three-dimensional matrix stiffness system is needed to clarify the interaction between matrix stiffness and tumor. Current studies have demonstrated that targeting the extracellular matrix is a potentially practical approach to treating tumors. Unfortunately, recent drug studies targeting extracellular matrix are limited to cellular and animal studies and need more detailed molecular mechanism studies, clinical specificity, and safety testing. Matrix stiffness promotes tumor progression and demonstrates potential as a target for antitumor therapy. In-depth studies on molecular mechanisms of matrix stiffness and tumors and the design of novel drugs targeting matrix stiffness will benefit tumor patients in the future.

Fig. 3. The molecular network of matrix stiffness regulating tumor progression(By Figdraw). Matrix stiffness can promote tumor progression by upregulating and downregulating various mechanically sensitive molecules. |
Ethics approval and consent to participate
Not applicable.
Conflict of interest
The authors declare that they have no conflict of interest.
Author contributions
RL and GS designed the review. RL drafted the manuscript, GS revised the review. All authors approved of the manuscript.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgments
This work was supported by grants from the National Natural Science Foundation of China (11832008), the Fundamental Research Funds for the Central Universities (2023CDJXY-051) and the Open Foundation of Guangdong Provincial Key Laboratory of Systems Biology and Synthetic Biology for Urogenital Tumors (2021B0303010151).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}