
1. Introduction
Coronavirus disease 19 (COVID-19) is the ongoing pandemic affecting millions of lives globally. The disease is caused by a highly infectious polycistronic (+) single-stranded retrovirus i.e., the SARS-CoV-2. Greater infectivity, transmissibility, and pathogenicity of SARS-CoV-2 compared to other SARS-CoVs might be due to the presence of a polybasic furin cleavage site at the S1/S2 junction of spike glycoprotein [1]. It invades various systems of the body [2,3,4]. The binding of spike glycoprotein with its cognate receptor angiotensin-converting enzyme 2 (ACE2) is followed by serine protease or cathepsin-mediated cleavage at S1/S2 junction [5]. This exposes a furin cleavage site at the S2. Furin cleavage site is targeted by serine protease to release a fusion peptide which plays critical role in viral-host membrane fusion and entry of the viral genome into the host cell [6]. Several mutations in a short period have been detected in the spike protein of the virus, and such mutations have produced several “variants of concern” viz. alpha, beta (B.1.351), gamma (P.1), delta (B.1.617.2), and omicron (B.1.1.529) [7]. Severity of the disease was higher in beta, gamma, and delta variants compared to the initial variant of SARS-CoV-2 whereas the impact of omicron is still unclear [8]. Notably, the high mutation rate in spike protein of SARS CoV-2 raises serious concern regarding the effectiveness of vaccines and the uncertainty of the current pandemic in the future. Hence repurposing of old drugs and the development of modified vaccines as well as therapeutic anti-viral agents are indispensable in the current situation. In this case, phytochemicals might play an important role in fighting COVID-19 infections on a broader scale worldwide.
Many phytochemicals have been assessed for their inhibitory effects on viral proteins and entry factors in host cells to restrain SARS-CoV-2 infection. Among several potential anti-viral phytochemicals, phenolic compounds from various plant sources could be an area of interest. For instance, polyphenol luteolin has been reported to bind spike protein of SARS-CoV with high affinity to impair its interaction with the ACE2 and subsequent invasion in to the host cells [9]. Another flavonoid naringin was reported to interact with the viral spike protein with a binding energy of −9.8 kcal/mol [10]. Glycosylated flavonoids could have greater inhibitory impacts against SARS-CoV-2 proteases. For example, Quercetin-3-O-rhamnoside was documented to block SARS-CoV-2 main protease (Mpro) with the highest binding affinity of −9,7 kcal/mol [11]. Quercetin derivatives such as quercetin 3-O-arabinoside 7-O-rhamnoside and quercetin-3-neohesperidoside-7-rhamnoside have been reported as potent inhibitors of spike glycoprotein, papain-like protease, and 3C-like protease of SARS-CoV-2 [12]. Six flavonoids such as eupatilin, 3′,4′-dimethoxyluteolin, 5,7,3′-trihydroxy-6,4′,5′-trimethoxyflavone, hispidulin, apigenin, and velutin were isolated from the areal part of Artemisia sublessingiana, and their binding affinity was tasted for Mpro. These polyphenols exhibited promising binding modes with free energies ranging from −6.39 to −6.81 (kcal/mol). Thus the implication of phytochemicals, particularly the phenolic compounds, might be helpful at least to some extent, in prophylaxis and treatment of COVID-19.
Green tea is one of the most extensively consumed beverages in the world. It is known to have multiple physiological as well as pharmacological health benefits. For instance, green tea has the potency to reduce the risk of cancer, cardiovascular ailments, and liver disease [13]. In addition, antioxidative, anti-inflammatory, antiviral, antiarthritic, antibacterial, and neuroprotective effects have also been reported [14,15,16,17]. Interestingly, most of the medicinal properties of green tea are attributed to various polyphenols, mainly flavanols and flavonols, which represent almost 30% of fresh leaf dry weight [18].
Among several phenolic compounds found in green tea, epicatechin gallate (ECG) is an important one (Fig. 1). It is a flavan-3-ol type of polyphenol having strong anti-oxidative potential. Apart from this, ECG has been reported to suppress the activities of pro-inflammatory mediators such as PGE2, TNF-α, IL-1β, and IL-6 [19]. Sometimes pharmacological benefits like anti-tumorigenic effects of ECG are superior to epigallocatechin gallate (EGCG), which is best studied by catechin and the major polyphenol present in green tea [20]. Studies involving the anti-viral activities of EGCG are plentiful, and they have been reported to block SARS-CoV-2 infection by inhibiting spike protein binding to ACE2 receptor [21]. However studies exploring the anti-SARS CoV-2 potential of ECG are rare. Hence, the present study aims to investigate the anti-SARS-CoV-2 efficacy of ECG using an in silico approach. The binding affinity of ECG for several entry factors, such as angiotensin converting enzyme 2 (ACE2) receptor, neuropilin, furin, and cathepsin L has been investigated in this study. These entry factors allow virions of SARS-CoV-2 to interact with the host's cell membrane for sub-cellular internalization of the viral genome. Moreover, the present in silico study also aimed to explore the binding affinity of ECG for several proteases and viral non-structural proteins (NSPs) that assist in the replication and proliferation of SARS-CoV-2 within the host cell.
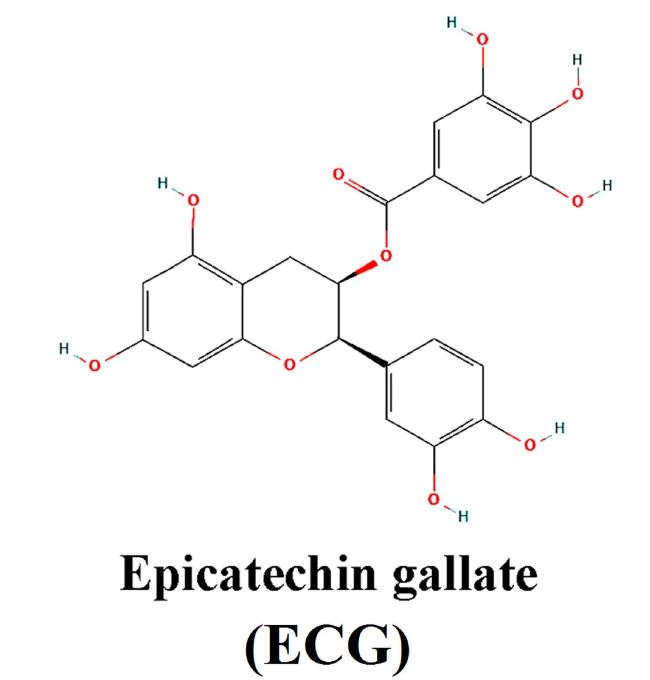
Fig. 1. Chemical structure of ECG. |
2. Materials and methods
2.1. Structure retrieval and preparation of ligand
Three-dimensional structure of the ligand ECG was retrieved from PubChem with CID 107905 (https://pubchem.ncbi.nlm.nih.gov/compound/Epicatechin-gallate). The molecular formula of the compound is C22H18O10. IUPAC name of the ligand is registered as [(2R,3R)-2-(3,4-dihydroxyphenyl)-5,7-dihydroxy-3,4-dihydro-2H-chromen-3-yl]3,4,5-trihydroxybenzoate. Chemical structure of EGC was processed using AutoDock 4.2 tool and saved as ligand.pdbqt format file for docking analysis.
2.2. Structure retrieval and preparation of SARS-CoV-2 proteins and entry factors
SARS-CoV-2 proteins and entry factors were retrieved from Protein Data Bank (PDB) as.pdb files (https://www.rcsb.org/). Three-dimensional structures of SARS-CoV-2 entry factors like ACE2, Neuropilin, Furin, and Cathepsin L (CTSL) with IDs 7zdq, 4deq, 7qxy, and 3 hha, respectively, were retrieved from PDB. Similarly, three-dimensional structures of eight SARS-CoV-2 proteins such as spike protein receptor binding domain (RBD), main protease (MPro), papain-like protease (PLPro), RNA-dependent RNA Polymerase (RdRP), Replication transcription (RepTrans) complex, NSP7-NSP8 complex, NSP13, NSP14, NSP15, and NSP16 with IDs 7lm9, 7nwx, 6w9c, 6m71, 7cyq, 6yhu, 6jyt, 7qif, 6w01, and 7JIB respectively were retrieved from PDB. These files were used by AutoDock 4.2 for the removal of water molecules, homogenous distribution of charges, and deletion of native ligands from the binding pockets to minimize the binding energy of the ECG during the docking analysis.
2.3. Docking procedure
Prior to the initiation of molecular docking, SARS-CoV-2 proteins and entry factors were processed using AutoDock 4.2. For this purpose, native ligands were removed from the binding pockets of concerned proteins. At the same time, the amino acids of the binding pockets were identified so that the grid box could be placed at the binding pockets to designate the docking site for ECG. In case of proteins where native ligands were not present, a grid box was placed to cover the entire protein wherever possible. Water molecules from the proteins were removed to facilitate the docking of polyphenols by minimizing the hindrance imposed by water molecules. Kollman electronic charges were homogenously applied to all atoms of proteins to get better binding affinity. Finally, molecular docking was conducted using a command prompt and AutoDock Vina [22,23,24,25]. Output files of the molecular docking were ligand_out.pdbqt and log.txt files. Log.txt file contained the binding affinities. Ligand_out.pdbqt files were opened using Chimera X to visualize the protein-ECG binding and topology. Later, LIGPLOT+ was used to identify amino acids that formed Hydrogen (H) bonds and established hydrophobic interactions with the ECG.
3. Results
3.1. Binding of ECG with SARS-CoV-2 entry factors
3.1.1. ACE2
The present in silico study has revealed the binding potential of ECG with the ACE2 receptor. Lowest binding energy for the interaction was recorded as −8.3 kcal/mol. ECG formed H-bonds with the amino acids Tyr196; Arg219; Ala396. Hydrophobic interactions were established between Gln102; Gly205, Gln98, Asp206; Glu208; Asn210; Leu95; Lys562; and Asn397 amino acids of ACE2 and ECG.
3.1.2. Neuropilin
ECG interacted with neuropilin with a binding energy of −8.0 kcal/mol. Several amino acids of the protein were engaged in H-bonds and other non-covalent interactions. Gln17 established H-bond. Other non-covalent interactions were made between Asp56, Arg50, Ser21, Arg129, Asp16, Trp52, and Lys134 of neuropilin and ECG.
3.1.3. Furin
ECG interacted with furin with several interactions. H-bonds were produced by Ile312; Gln488; Ser311; Ala532, Asp264, Pro266; and Pro266 of the enzyme. Hydrophobic interactions were established between Phe275; Glu271; Arg490; Asp530; Asn310; and Trp531 of furin and ECG. Lowest binding energy was recorded as −8.9 kcal/mol (Fig. 2).
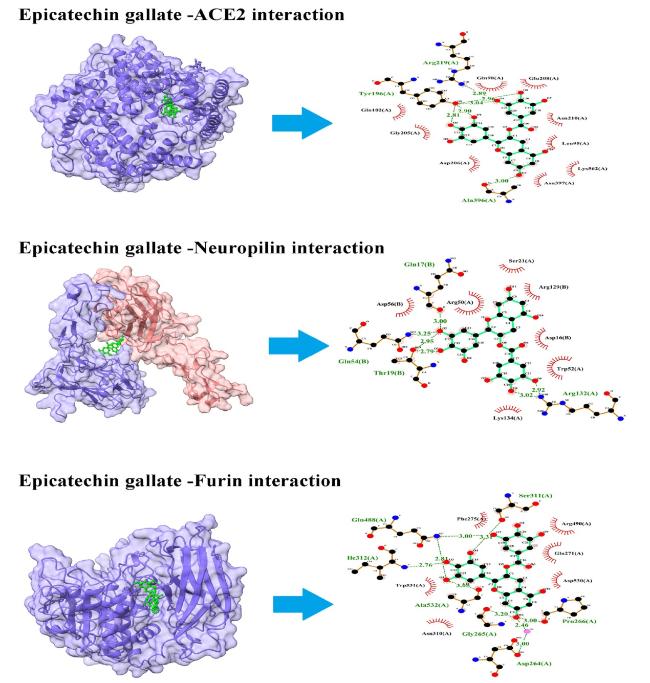
Fig. 2. Interaction of ECG with entry factors. ECG blocks several amino acid residues of ACE2, neuropilin, and furin at binding pockets via H- bonds and other non-covalent interactions. Left panel shows the site of protein-ligand interactions. Right panel depicts the amino acid residues of concerned protein involved in H-bonds and hydrophobic interactions with ECG. |
3.1.4. CTSL
ECG showed the highest binding affinity with Cathepsin L with an affinity score of −10.2 kcal/mol. Several amino acids of the protein were blocked by non-covalent interactions. Amino acids Lys124; Asp155; Arg205; and Tyr217 were involved in H-bond formation whereas amino acids Val113(B); Phe152(A); Cys152(A); Thr219(B); His208(A); Arg206(A); Aer158(A); Arg40(B); Val220(B); Glu159(A); Val129(B); Ser157(A); Thr128(B); and Cys156(A) established non-covalent hydrophobic interactions with ECG (Fig. 3).
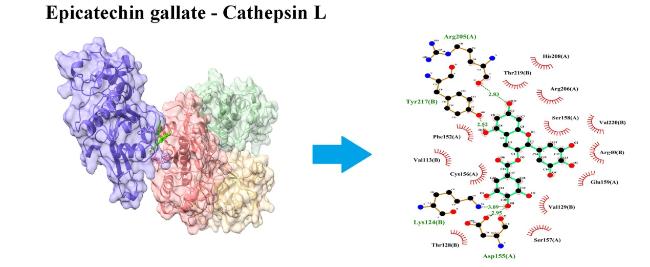
Fig. 3. Interaction of ECG with cathepsin L (CTSL). ECG blocks several amino acid residues of CTSL at binding pockets via H- bonds and other non-covalent interactions. Left panel shows the site of protein-ligand interactions. Right panel depicts the amino acid residues of CTSL involved in H-bonds and hydrophobic interactions with ECG. |
3.2. Binding of ECG with SARS-CoV-2 proteins
3.2.1. Spike protein RBD
RBD of Spike protein is crucial for sub-cellular invasion of the virus. ECG was noticed to interact with the RBD of spike protein with a binding energy of −7.4 kcal/mol. ECG blocked the amino acids viz. Asp415; Leu503; Phe501; Met417; and Pro450 of RBD with non-covalent bonds. Other amino acids such as Glu502; Ser500; Phe451; and Pro413 of RBD were blocked by ECG through stable hydrophobic interactions.
3.2.2. Main protease (Mpro)/NSP5
The present in silico study has unraveled the binding potential of ECG with the NSP5. The best binding energy for the interaction was recorded as −8.4 kcal/mol. ECG formed H-bonds with the five amino acids viz. His41; Cys145; His163; Phe140; and Thr190. Hydrophobic interactions were established between Leu141; Glu166; Met165; Gln189; and Pro168 of NSP5 and ECG.
3.2.3. Papain-like protease (PLpro)
ECG interacted with PLpro with a binding energy of −8.9 kcal/mol. Several amino acids of the protein were engaged in H-bonds and non-covalent interactions. Strong H-bonds were formed by Asp108; Asn109; Thr158; Glu161; Val159; and Gln269. Non-covalent interactions were developed between Glu161; Leu162; Asn109; Val159; Gly160; Leu162; Gln269; and Gln269 of PLpro and ECG (Fig. 4).

Fig. 4. Interaction of ECG with viral spike protein and proteases. ECG blocks several amino acid residues of receptor binding domain (RBD) of spike protein, main protease (Mpro), and papain like protease (PLpro) at binding pockets via H- bonds and other non-covalent interactions. Left panel indicates the site of protein-ligand interactions. Right panel represents the amino acid residues of concerned protein involved in H-bonds and hydrophobic interactions with ECG. |
3.2.4. RNA-dependent RNA polymerase (RdRP)/NSP12
ECG interacted with RdRP with several interactions. H-bonds were formed by four amino acids viz. His133; Asn781; Ser784; and Ser709 of the enzyme. Hydrophobic interactions were established between Thr710; Ala706; Lys780; Tyr129; Asp135; Asn138; Tyr32; Lys47 of RdRP and ECG. Lowest binding energy was recorded as −8.6 kcal/mol.
3.2.5. RepTrans complex
RepTrans complex is vital for the replication of the virus in a sub-cellular environment. ECG was noticed to interact with the RepTrans complex with a binding energy of −9.0 kcal/mol. ECG blocked ten amino acids viz. Asn781; Ser784; Tyr129; Ser784; Tyr32; Lys47; Ser709; Lys780; His133; and Phe134 of RepTrans complex with stable non-covalent bonds. Other amino acids such as Gly774; Aer778; Ala771; Thr710; Asp711; Ala46; and Asp135 of RepTrans complex were engaged with ECG through hydrophobic interactions (Fig. 5).
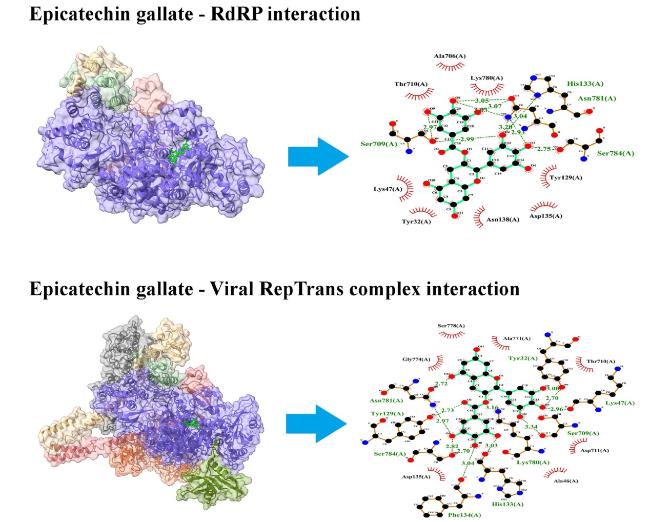
Fig. 5. Interaction of ECG with viral RdRP and RepTrans complex. ECG blocks several amino acid residues of RdRP and RepTrans complex at binding pockets via H-bonds and non-covalent interactions. Left panel shows the site of protein-ligand interactions. Right panel depicts the amino acid residues of concerned protein involved in H-bonds and hydrophobic interactions with ECG. |
3.2.6. NSP7-NSP8 complex
ECG interacted with NSP7-NSP8 complex with a binding energy of −8.3 kcal/mol. Several amino acids of the protein were engaged in H-bonds and non-covalent interactions. H-bonds were formed by Ser15; Gln18; and Glu19. Non-covalent interactions were developed between Met87; Arg21; Gln18; Leu14; Arg80; His36; Asn37; and Val83 of NSP7-NSP8 complex and ECG.
3.2.7. NSP13
NSP13 is crucial for the replication of SARS-CoV-2 in the sub-cellular moiety. ECG was noticed to interact with the NSP13 with the lowest binding energy of −8.7 kcal/mol. ECG blocked four amino acids viz. Ala4; Glu136; Leu235; and Arg129 of RBD with strong H-bonds. Other amino acids such as Arg21(A); Pro23(A); Phe24(A); Gly3(A); Leu32(A); Val6(A); Pro234(A); and Phe133(A) of NSP13 were blocked by ECG through hydrophobic interactions (Fig. 6).

Fig. 6. Interaction of ECG with viral NSP7-NSP8 complex and NSP13. ECG blocks several amino acid residues of NSP7-NSP8 complex and NSP13 at binding pockets via H-bonds and other non-covalent interactions. In the left panel, sites of protein-ligand binding have been presented. Amino acid residues of concerned protein involved in H-bonds and hydrophobic interactions with ECG have been presented in right panel. |
3.2.8. NSP14
ECG interacted with NSP14 with several interactions. H-bonds were produced by Gln313; Asn386; and Asn388 of the enzyme. At NSP14-ECG interface, hydrophobic interactions were established by Trp385; Asn306; Phe426; Phe401; Phe506; Thr428; Pro355; and Arg310 amino acid residues of NSP14. Least binding energy was recorded as −9.1 kcal/mol.
3.2.9. NSP15
ECG interacted with NSP15 with a binding energy of −7.9 kcal/mol. Several amino acids of the protein were engaged in non-covalent interactions. H-bonds were formed by Glu234; Phe241; Gly239; and Arg258. Protein ligand binding was further strengthened by Met219; Tyr238; Asp240; Ser242; Thr286; Glu261; and Lys257 residues of NSP15 through hydrophobic interactions.
3.2.10. NSP16
The present in silico study has unraveled the binding potential of ECG with the NSP16. The best binding energy for the interaction was recorded as −8.5 kcal/mol. ECG formed H-bonds with the six amino acids viz. Tyr132; Asn29; Asn198; Lys170; Glu173; and Ser202 of NSP16. Hydrophobic interactions were established between Glu203; Ser201; His138; Leu27; Val139; Cys25; His174; and Thr172 amino acids of NSP16 and ECG (Fig. 7).
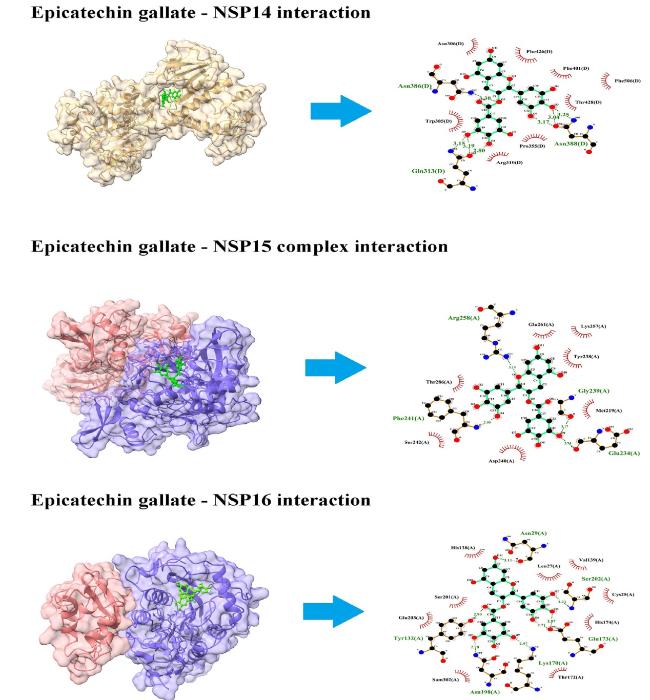
Fig. 7. Interaction of ECG with viral NSP14, NSP15, and NSP16. ECG blocks several amino acid residues of NSP14, NSP15, and NSP16 at binding pockets via H-bonds and non-covalent interactions. Left panel shows the site of protein-ligand interactions. Right panel depicts the amino acid residues of concerned protein involved in H-bonds and hydrophobic interactions with ECG. |
4. Discussion
Table 1. Binding affinity of protein-ligand complex. |
| Protein ligand complex | Binding affinity (kcal/mol) |
|---|---|
| ECG ∼ ACE-2 | −8.3 |
| ECG ∼ Neuropilin | −8.0 |
| ECG ∼ Furin | −8.9 |
| ECG ∼ Cathepsin L | −10.2 |
| ECG ∼ spike protein RBD | −7.4 |
| ECG ∼ COVID-19 Mpro/NSP5 | −8.4 |
| ECG ∼ PLpro | −8.9 |
| ECG ∼ RdRP/NSP12 | −8.6 |
| ECG ∼ RepTrans complex | −9.0 |
| ECG ∼ NSP7-NSP8 complex | −8.3 |
| ECG ∼ NSP13 | −8.7 |
| ECG ∼ NSP14 | −9.1 |
| ECG ∼ NSP15 | −7.9 |
| ECG ∼ NSP16 | −8.5 |
Table 2. Hydrogen bonds and Hydrophobic interactions established between ECG and amino acid residues of host entry factors/viral proteins of SARS-CoV-2. |
| Protein ligand complex | Amino acids of proteins establishing Hydrogen bonds with ligand | Amino acids of proteins establishing hydrophobic interactions with ligand |
|---|---|---|
| ECG ∼ ACE-2 | Tyr196; Arg219; Ala396 | Gln102; Gly205, Gln98, Asp206; Glu208, Asn210, Leu95, Lys562, Asn397 |
| ECG ∼ Neuropilin | Gln17; Gln54, Thr19, Arg132 | Asp56, Arg50, Ser21, Arg129, Asp16, Trp52, Lys134 |
| ECG ∼ Furin | Ile312; Gln488; Ser311; Ala532, Asp264, Pro266; Pro266 | Phe275; Glu271; Arg490; Asp530; Asn310; Trp531 |
| ECG ∼ Cathepsin L | Lys124; Asp155; Arg205; Tyr217 | Val113(B); Phe152(A); Cys152(A); Thr219(B); His208(A); Arg206(A); Aer158(A); Arg40(B); Val220(B); Glu159(A); Val129(B); Ser157(A); Thr128(B); Cys156(A) |
| ECG ∼ spike protein RBD | Asp415; Leu503; Phe501; Met417; Pro450 | Glu502; Ser500; Phe451; Pro413 |
| ECG ∼ Mpro/NSP5 | His41; Cys145; His163; Phe140; Thr190 | Leu141; Glu166; Met165; Gln189; Pro168 |
| ECG ∼ PLpro | Asp108; Asn109; Thr158; Glu161; Val159; Gln269 | Glu161; Leu162; Asn109; Val159; Gly160; Leu162; Gln269; Gln269 |
| ECG ∼ RdRP/NSP12 | His133; Asn781; Ser784; Ser709 | Thr710; Ala706; Lys780; Tyr129; Asp135; Asn138; Tyr32; Lys47 |
| ECG ∼ RepTrans complex | Asn781; Ser784; Tyr129; Ser784; Tyr32; Lys47; Ser709; Lys780; His133; Phe134 | Gly774; Aer778; Ala771; Thr710; Asp711; Ala46; Asp135 |
| ECG ∼ NSP7-NSP8 complex | Ser15; Gln18; Glu19 | Met87; Arg21; Gln18; Leu14; Arg80; His36; Asn37; Val83 |
| ECG ∼ NSP13 | Ala4; Glu136; Leu235; Arg129 | Arg21(A); Pro23(A); Phe24(A); Gly3(A); Leu32(A); Val6(A); Pro234(A); Phe133(A) |
| ECG ∼ NSP14 | Gln313; Asn386; Asn388 | Trp385; Asn306; Phe426; Phe401; Phe506; Thr428; Pro355; Arg310 |
| ECG ∼ NSP15 | Glu234; Phe241; Gly239; Arg258 | Met219; Tyr238; Asp240; Ser242; Thr286; Glu261; Lys257 |
| ECG ∼ NSP16 | Tyr132; Asn29; Asn198; Lys170; Glu173; Ser202 | Glu203; Ser201; His138; Leu27; Val139; Cys25; His174; Thr172 |
ACE2 is an entry receptor for SARS-CoV-2. Spike glycoprotein of the virion is composed of a trimeric head seated on the top of a trimeric stalk. The trimeric head bears a receptor binding domain (RBD). It binds with the ACE2 for internalization of virions into the host cell. This is a critical step in the incorporation of the viral genome into the host cell. In the present study, ECG was found to bind with ACE2 with higher binding affinity. Three H-bonds and several hydrophobic interactions were detected at the ECG-ACE2 interface. Our finding was in line with another in silico study where kobophenol A, a tetramer of resveratrol, interacted with ACE2 hydrophobic pocket [26]. Similarly, three compounds, viz. deslanoside, digitoxin, and digoxin, were reported to show strong binding with ACE2 at the H1-H2 binding pocket [27]. The free energies of Luteolin and Chrysin were also analyzed through the Molecular Mechanics/Poisson-Boltzmann Surface Area method (MM/PBSA), which confirmed effective binding to ACE2 [28].
Neuropilin, which regulates axon guidance, angiogenesis, and vascular permeability, also acts as a host factor for SARS-CoV-2 infection. It is abundantly expressed in the olfactory and respiratory epithelium, with highest magnitude of expression in endothelial and epithelial cells. Neuropilin, when co-expressed with ACE2 and TMPRSS2 can bind the cleaved S1, thereby enhancing infection [29]. Blocking of neuropilin by antibody-like mAb3 can reduce the infection [29]. Thus neuropilin inhibitors might restrict SARS-CoV-2 entry into the host cells. In the present study, ECG binds with neuropilin with various non-covalent bonds and interactions. Similar to this, Quercetin derivates like Quercetin-3-O-(2″,6″-digalloyl)- β-D-galactopyranoside, Quercetin-3-O-[(6-O-feruloyl)-β-D-glucopyranosyl- (1 → 2)-β-Dgalactopyranoside] and Quercetin-3-O-[β-D-glucopyranosyl-(1 → 2)- β-D-galactopyrano-side] are reported as stronger binders for neuropilin [30]. Such binding by phytocompound might restrict interaction between cleaved S1 and neuropilin which could reduce viral entry into the host cells.
The spike protein of SARS-CoV-2 is initially processed at S1/S2 junction through host cell protease. Such priming results in the incorporation of four critical amino acids (PRRA) at S1/S2 interface, leading to the formation of a canonical PRRAR685↓ furin-like cleavage site [31]. Studies involving cell culture and animal models have indicated the undeniable implication of furin in the priming of S1/S2 interface [32,33,34]. Therefore inhibition of furin could potentially abolish SARS-CoV-2 entry into the host cell. In the present in silico study, ECG blocked several amino acid residues of furin through H-bonds and hydrophobic interactions. Similar to the present finding, some phytocompounds like quercetin-3-O-α-L-rhamnopyranoside, kaempferol 3-O-α-L-rhamnopyranosyl-(1 → 4)-β-D-glucopyranoside, and isovitexin have also been shown to exhibit high binding affinity for furin [35]. Lau et al. [36] have reported that virions having a mutation at S1/S2 sites replicate weakly in tracheal and lung tissues compared to the wild type. Therefore, ECG could be a prophylactic agent for SARS-CoV-2 agent.
CTSL is a member of lysosomal cysteine protease having an L domain of α-helix and an R domain of β-sheet. CTSL exhibits endo- or exopeptidase activities. Fusion of SARS coronavirus with the host plasma membrane relies upon the proteolytic cleavage of spike protein by host CTSL [37]. Thus, proteolytic action of CTSL is required to augment SARS-CoV-2 infection. In the present study, amino acid residues of CTSL were blocked by ECG via many non-covalent bonds and interactions. Similar to this finding, a group of flavonoids have also been observed to inhibit five molecular targets of SARS-CoV-2, including CTSL [38]. Vivek-Ananth et al. [39] analyzed potential natural product inhibitors of human proteases, which are key to SARS-CoV-2 infection. They identified ararobinol, (+)-oxoturkiyenine, and 3α,17α-cinchophylline as potential inhibitors of CTSL due to their high binding affinity. Thus, CTSL inhibitors could play a crucial role in SARS-CoV-2 infection.
ECG interacted with RBD of spike protein with H-bonds and several hydrophobic interactions. Similar to this finding, another flavonoid naringin was found to interact with the spike protein through H-bonds and several hydrophobic interactions. Authors concluded that naringin might act as a potent viral protein inhibitor [10]. Kobophenol A has also been shown to block S1-RBD thereby inhibiting SARS-CoV-2 binding with ACE2 [26]. In silico study involving withanone, a natural product found in Withania somnifera has demonstrated that, the chemical can efficiently bind at the interacting interface of the ACE2-RBD complex to energetically destabilize it. This finding was further validated using humanized zebrafish model [40]. In an in vitro RBD-ACE2 binding assay, three compounds such as heparin sodium, aurintricarboxylic acid, and ellagic acid, were found to exert an effective binding inhibition for which molecular docking analysis was conducted for potential binding sites for these molecules in spike RBD [41].
Upon entry into the host cytoplasm, RNA genome of SARS-CoV-2 directly attaches with the host ribosome for translation into two large polyproteins (pp1a and pp1b). These polyproteins undergo proteolytic cleavage by proteases viz. Mpro and CLpro, which are needed for the packaging of new virions. Mpro is considered as virus’ “Achilles Heel” and therefore, it is an attractive drug target to inhibit viral replication within the host cell. In the present study, ECG blocked the binding pocket of Mpro via several non-covalent interactions. Similar to the present finding, some phytocompounds like quercetin-3-O-α-L-rhamnopyranoside, kaempferol 3-O-α-L-rhamnopyranosyl-(1 → 4)-β-D-glucopyranoside, and isovitexin have been shown to exhibit a high binding affinity for SARS-CoV-2 Mpro [35]. A molecular dynamics study has indicated the binding mode of quercetin with Mpro via many amino acid interactions [42]. Several other flavonoids like fisetin, apigenin, asquercetin, kaemferol, myricetin, and daidzein have also been reported to inhibit Mpro by disrupting the activation/dimerization of the proteases [43]. In our study, ECG interacted with both Mpro and PLpro with higher affinity. Notably, it has been suggested that combined inhibition of Mpro and PLpro as also observed in the present study, is more effective in blocking infection by SARS-CoV-2 variants [43].
PLpro, with high content of cysteine residues, is an essential enzyme for SARS-CoV-2 replication. It is involved in the processing of coronavirus polyproteins in order to produce functional replicase complex and to speed up viral replication [44]. It is also implicated in proteinous post-translational modifications via deubiquitinating and deISGylating effects that are required to evade host's defense mechanism [45]. In the present study, ECG has been observed to block many amino acid residues of PLpro through several non-covalent bonds and interactions that might impede the functional attributes of the enzyme. Anti-SARS-CoV-2 activity was exhibited by metergoline, which could inhibit PLpro activity to block sub-cellular entry of virions [46]. Another compound viz. rac5c develops essential interactions (H-bond and hydrophobic interactions) with PLpro as validated by another in silico study [47].
The core replication machinery of SARS-CoV-2 mainly constitutes the NSP12, also known as the major RNA-dependent RNA polymerase (RdRp) and NSP7-NSP8 complex as co-factors. Polymerase activity of NSP12 is greatly enhanced by NSP7-NSP8 [48]. Though some other NSP subunits are required to complete the full repertoire of replication and transcription, NSP12-NSP7-NSP8 represents the minimal complex for polymerization of viral genome. Corilagin is a phytocompound that is shown to bind directly to RdRp to block its polymerase activity [49]. Moreover computational modeling has predicted that corilagin interacts with the palm domain of RdRp to prevent the conformational change required for nucleotide incorporation by RdRp [49]. In a cell-based assay, 2-((1H-indol-3-yl)thio)-N-phenyl-acetamides specifically targeted RdRp and inhibited viral replication in a dose-dependent manner [50]. In silico study showed that silibinin possesses the binding affinity for S protein and RdRP as −7.78 and −7.15 kcal/mol, respectively. IC50 of silibinin for S protein, Mpro, and RdRP was reported as 0.029, 0.021, and 0.042 μM, respectively [51]. Comparative molecular docking revealed the binding affinity of silibinin for S protein and RdRP as −7.78 and −7.15 kcal/mol, respectively. Plant-derived non-nucleoside analogues bind to the allosteric sites of RdRP complex (nsp7/nsp8/nsp12) to change the redox state and to alter antiviral activity [52]. Similar to these studies, ECG interacted with the RdRP complex via non-covalent interactions that could subvert the replicative potential of SRAS-CoV-2.
NSP13 is active as NTPase and RNA helicase. It stimulates RdRp complex for the insertion of ribonucleotides during viral replication [53]. NSP13 also suppresses type I IFN production and subsequent signaling of anti-viral defense system [54]. Thus, the involvement of NSP13 is critical in SARS-CoV-2 infection and pathogenicity. Studies have documented that inhibiting NSP13 with bismuth salts and other agents like clofazimine blocks viral replication [55,56]. Natural compounds like myricetin, quercetin, kaempferol, and flavanone were reported to inhibit the unwinding activity of NSP13 at nanomolar concentrations [57]. Moreover, computational studies confirmed a unique pattern of interactions between amino acid residues of NSP13 and the natural compounds. Other naturally occurring flavonoids like myricetin and scutellarein have also been demonstrated to inhibit the helicase activity of NSP13 in vitro by affecting the ATPase activity [58]. Present in silico study has unveiled that ECG can bind with NSP13 with high affinity by establishing many H-bonds and hydrophobic interactions that could block the helicase activity of NSP13 during SARS-CoV-2 replication.
NSP14 is the core component aiding to viral replication and pathogenesis. Its N-terminal domain has endoribonuclease (ExoN) activity which participates in the proofreading of viral RNA during replication. The C terminal domain of NSP14 has N7-methyltransferase (N7-MTase) activity that helps in 5’ cap methylation of viral RNA to enable viral evasion of the host defense system [59]. Thus, inhibition of NSP14 could be a promising strategy to abort the proofreading of the viral genome. In the present study, ECG blocked several amino acid residues of NSP14 that could prevent NSP14 function. In a study, N7-Methyltransferase activities of SARS-CoV-2 NSP14 were inhibited by certain sulfonamide-based bisubstrate analogues [60]. Moreover, molecular docking analysis supported the structure-activity relationships of these analogues. Another in silico study investigated the binding potential of the drug ritonavir to the active site of NSP14 and found significant binding affinity. Authors of this study concluded that ritonavir may act as an effective inhibitor of the NSP14 protein [61].
NSP15 is an endoribonuclease commonly called endoU and is characteristic of all coronavirus family members. It preferentially cleaves RNA substrates 3′ of uridines. Endonuclease function of NSP15 is critical for life cycle of coronavirus. Moreover, it is important to evade host's innate immune defense system [62]. Loss of NSP15 endoribonuclease activity in porcine epidemic diarrhea coronavirus (PEDV) is associated with the activation of IFN response and reduction in viral shedding and mortality in infected piglets [62]. Thus blocking NSP15 activity could be a potential therapeutic agent against SARS-CoV-2 infection. Crystallography, biochemical, and whole-cell assays have revealed that the drug tipiracil interacts with uridine binding pocket in the active site of NSP15 to inhibit the endoribonuclease activity [63]. In an in vitro study, three phytochemicals viz. epigallocatechin gallate, baicalin, and quercetin, were identified as potential inhibitors of NSP15 endoribonuclease activity [64]. Notably, epigallocatechin gallate effectively neutralized SARS-CoV-2 strains in plaque reduction neutralization tests. In the present study, ECG blocked NSP15 through H-bonds and several hydrophobic interactions that could potentially reduce viral replication and enhance the anti-viral defense system within the host.
NSP16 is an important component during the coronavirus replication cycle. It is actively involved in the capping of transcribed viral mRNA. NSP16 transfers a methyl group from its S-adenosylmethionine cofactor to the 2′ hydroxyl of ribose sugar of viral mRNA [65]. This methylation improves the translation process and camouflages the viral mRNA so that it is not recognized by intracellular pathogen recognition receptors such as IFIT and RIG-I [66]. Therefore, it is also crucial to evade host's anti-viral defense system. It is important to note that inhibiting or knocking out 2′-O-MTase activity severely attenuates coronavirus replication and its infectivity [67]. Thus, screening agents that can block NSP16 activity is a promising therapeutic strategy. In an in silico study, four compounds of zinc database exhibited better docking results compared to Sinefungin for NSP16 [68], suggesting a potential role of these compounds in reducing SARS-CoV-2 infection. In the present study, ECG has blocked the amino acid residues of NSP16 that could prevent the methyl transfer activity of the enzyme resulting in reduced viral replication (Fig. 8). Like microbial infections, other xenobiotics could also be detrimental to the health [69,70,71,72,73,74,75,76,77,78,79,80]. Phytochemicals could help mitigate such adverse effects [81,82]. Similarly, ECG might prevent infectious agents to replicate inside the body.

Fig. 8. Multi-target potency of ECG for SARS-CoV-2 entry and replication within the host cell. Spike protein of SARS-CoV-2 attaches with host ACE2 and undergoes proteolytic cleavage by host proteases viz. furin and neuropilin. This facilitates fusion and viral entry into the host cell. Cathepsin L (CTSL) helps in release of (+) ss viral RNA to the host cytoplasm. Viral genome uses host-translational machinery to generate viral polyproteins (pp1a, pp1b). pp1a generates viral proteases viz. Mpro and PLpro that cleaves pp1b into several non-structural proteins (NSPs). NSPs constitute the replication-transcription machinery for replication of viral genome and synthesis of (−) ss viral RNA is crucial for synthesis of structural proteins and their packing into new virions. These virions are then exocytosed out of the cells to infect the fresh one. ECG can bind with host entry factors, viral proteases, and NSPs to inhibit viral entry and replication within the host cell. |
Several similar compounds have been studied using molecular docking analysis to unravel their inhibitory potential against SARS-CoV-2. For instance, epigallocatechin gallate and gallocatechin-3-gallate have been found to interact with catalytic residues (His41 and Cys145) of Mpro with binding affinities ranging from −7.1 to −9.0 kcal/mol [83]. In another in silico study, epigallocatechin gallate was highly effective in inhibiting infection of live SARS-CoV-2 and human coronavirus (HCoV OC43). Epigallocatechin gallate mechanistically blocked the intermolecular interaction between ACE-2 and RBD of spike protein [84]. Kicker et al. [85] demonstrated the efficacy of green tea extract containing several catechins to neutralize the SARS-CoV-2. Molecular docking analyses of Camellia sinensis catechins have revealed that esterificating the 3'-OH as well as changing the configuration of C-3' or C-5' of catechins could help increase the efficacy against SARS-CoV-2 3CLpro [86]. With a binding score of −7.8 kcal/mol, epigallocatechin gallate has been observed to interact with the entry factor ACE2 [87]. In another study, the tea compounds epigallocatechin gallate and ECG showed virucidal activity against SARS-CoV-2, with IC50 values of 6.5 and 12.5 μM, respectively [88]. ECG has recently been reported to interact with inflammatory and pyroptosis-associated mediators of COVID-19 [89].Therefore, the binding potential of ECG to various entry and replication factors of SARS-CoV-2 could subvert virucidal activity.
Therefore the present study indicates the binding potential of ECG against various entry and replication factors of SARS-CoV-2. Nonetheless, results on binding potentials of ECG require further validation using cell-based and animal models. Hence, the findings of the present study open up futuristic testing (in vitro and in vivo) possibilities of ECG against COVID-19.
5. Conclusion
ECG showed high-affinity interactions with several SARS-CoV-2 entry factors, spike proteins, and NSPs that are critical to viral entry and replication into the host cells. Several amino acid residues in the binding pockets of these proteins were blocked by many non-covalent bonds and interactions formed by ECG. Such blockage could inhibit the obligatory interactions between viral spike protein and host cell receptors as well as proteases. This could potentially impede viral-host cell membrane fusion and transfer of viral genome into the host cytoplasm. NSPs are important constituents of the replication-transcription machinery of SARS-CoV-2 and hence inhibition of NSP activity could further reduce viral replication, m-RNA transcription, and translation of structural proteins within the host cells.
Ethical statement
Not applicable.
Author contribution
Prem Rajak: Conceptualization, Methodology, Investigation, Data Curation, Writing- Original draft preparation, Writing- Reviewing and Editing, Supervision. Abhratanu Ganguly: Writing- Original draft preparation, Data Curation.
Conflict of interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}